AOP ID and Title:
Graphical Representation
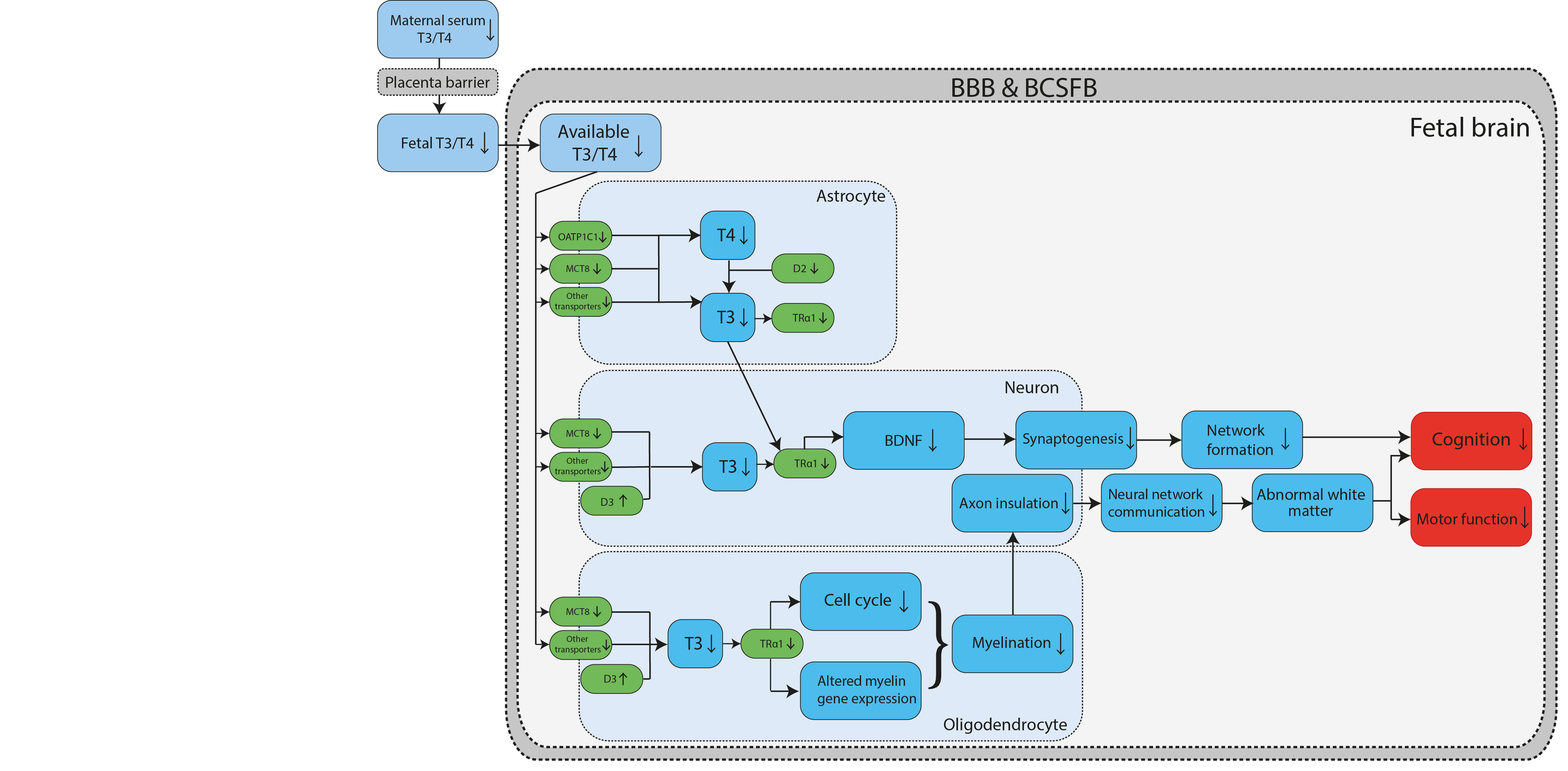
Status
| Author status | OECD status | OECD project | SAAOP status |
|---|---|---|---|
| Under development: Not open for comment. Do not cite |
Summary of the AOP
Events
Molecular Initiating Events (MIE), Key Events (KE), Adverse Outcomes (AO)
| Sequence | Type | Event ID | Title | Short name |
|---|---|---|---|---|
| MIE | 2258 | Inhibition, monocarboxylate transporter 8 (MCT8) | Inhibition, monocarboxylate transporter 8 (MCT8) | |
| MIE | 2376 | Inhibition, organic anion-transporting polypeptide 1C1 (OATP1C1) | Inhibition, OATP1C1 | |
| MIE | 1002 | Inhibition, Deiodinase 2 | Inhibition, Deiodinase 2 | |
| MIE | 1656 | Antagonism, Thyroid Receptor | TR Antagnoism | |
| KE | 2093 | Brain thyroid hormone (T4/T3) decreased | Decreased brain T4/T3 | |
| KE | 381 | Reduced levels of BDNF | BDNF, Reduced | |
| KE | 385 | Decrease of synaptogenesis | Synaptogenesis, Decreased | |
| KE | 1505 | Cell cycle, disrupted | Cell cycle, disrupted | |
| KE | 2105 | Decreased, myelin basic protein expression in oligodendrocytes | Decreased, mbp expression in oligodendrocytes | |
| KE | 2266 | Demyelination, increased | Demyelination, increased | |
| KE | 2378 | Impaired axonal insulation | Impaired axonal insulation | |
| KE | 2377 | Decrease, neural network maturation | neural network formation | |
| KE | 386 | Decrease of neuronal network function | Neuronal network function, Decreased | |
| KE | 2108 | Altered, white brain matter | Altered, white brain matter | |
| AO | 341 | Impairment, Learning and memory | Impairment, Learning and memory | |
| AO | 2231 | Locomotor function. Impaired | Impaired locomotor function |
Key Event Relationships
| Upstream Event | Relationship Type | Downstream Event | Evidence | Quantitative Understanding |
|---|---|---|---|---|
| Inhibition, monocarboxylate transporter 8 (MCT8) | adjacent | Brain thyroid hormone (T4/T3) decreased | Not Specified | Not Specified |
| Inhibition, organic anion-transporting polypeptide 1C1 (OATP1C1) | adjacent | Brain thyroid hormone (T4/T3) decreased | Not Specified | Not Specified |
| Inhibition, Deiodinase 2 | adjacent | Brain thyroid hormone (T4/T3) decreased | Not Specified | Not Specified |
| Brain thyroid hormone (T4/T3) decreased | adjacent | Antagonism, Thyroid Receptor | Not Specified | Not Specified |
| Antagonism, Thyroid Receptor | adjacent | Reduced levels of BDNF | Not Specified | Not Specified |
| Reduced levels of BDNF | adjacent | Decrease of synaptogenesis | Not Specified | Not Specified |
| Decrease of synaptogenesis | adjacent | Decrease, neural network maturation | Not Specified | Not Specified |
| Decrease, neural network maturation | adjacent | Locomotor function. Impaired | Not Specified | Not Specified |
| Decrease, neural network maturation | adjacent | Impairment, Learning and memory | Not Specified | Not Specified |
| Antagonism, Thyroid Receptor | adjacent | Cell cycle, disrupted | Not Specified | Not Specified |
| Antagonism, Thyroid Receptor | adjacent | Decreased, myelin basic protein expression in oligodendrocytes | Not Specified | Not Specified |
| Cell cycle, disrupted | adjacent | Demyelination, increased | Not Specified | Not Specified |
| Decreased, myelin basic protein expression in oligodendrocytes | adjacent | Demyelination, increased | Not Specified | Not Specified |
| Demyelination, increased | adjacent | Impaired axonal insulation | Not Specified | Not Specified |
| Impaired axonal insulation | adjacent | Decrease of neuronal network function | Not Specified | Not Specified |
| Decrease of neuronal network function | adjacent | Altered, white brain matter | Not Specified | Not Specified |
| Altered, white brain matter | adjacent | Impairment, Learning and memory | Not Specified | Not Specified |
| Altered, white brain matter | adjacent | Locomotor function. Impaired | Not Specified | Not Specified |
| Brain thyroid hormone (T4/T3) decreased | non-adjacent | Reduced levels of BDNF | Not Specified | Not Specified |
Overall Assessment of the AOP
References
Appendix 1
List of MIEs in this AOP
Event: 2258: Inhibition, monocarboxylate transporter 8 (MCT8)
Short Name: Inhibition, monocarboxylate transporter 8 (MCT8)
Key Event Component
| Process | Object | Action |
|---|---|---|
| thyroid hormone transport | monocarboxylate transporter 8 | decreased |
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:610 - Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | MolecularInitiatingEvent |
Biological Context
| Level of Biological Organization |
|---|
| Molecular |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| human | Homo sapiens | Moderate | NCBI |
| mouse | Mus musculus | Moderate | NCBI |
| zebrafish | Danio rerio | Moderate | NCBI |
| chicken | Gallus gallus | Moderate | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | Moderate |
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Event: 2376: Inhibition, organic anion-transporting polypeptide 1C1 (OATP1C1)
Short Name: Inhibition, OATP1C1
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:610 - Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | MolecularInitiatingEvent |
Biological Context
| Level of Biological Organization |
|---|
| Molecular |
Event: 1002: Inhibition, Deiodinase 2
Short Name: Inhibition, Deiodinase 2
Key Event Component
| Process | Object | Action |
|---|---|---|
| catalytic activity | type II iodothyronine deiodinase | decreased |
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:155 - Deiodinase 2 inhibition leading to increased mortality via reduced posterior swim bladder inflation | MolecularInitiatingEvent |
| Aop:156 - Deiodinase 2 inhibition leading to increased mortality via reduced anterior swim bladder inflation | MolecularInitiatingEvent |
| Aop:190 - Type II iodothyronine deiodinase (DIO2) inhibition leading to altered amphibian metamorphosis | MolecularInitiatingEvent |
| Aop:610 - Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | MolecularInitiatingEvent |
Stressors
| Name |
|---|
| iopanoic acid |
| PERFLUOROOCTANOIC ACID |
Biological Context
| Level of Biological Organization |
|---|
| Molecular |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| rat | Rattus norvegicus | Moderate | NCBI |
| human | Homo sapiens | High | NCBI |
| pigs | Sus scrofa | Moderate | NCBI |
| Oreochromis niloticus | Oreochromis niloticus | Moderate | NCBI |
| zebrafish | Danio rerio | Moderate | NCBI |
| fathead minnow | Pimephales promelas | Moderate | NCBI |
| African clawed frog | Xenopus laevis | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | Moderate |
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Taxonomic: Deiodination by DIO enzymes is known to exist in a wide range of vertebrates and invertebrates. This KE is plausibly applicable across vertebrates. Reports of inhibition of DIO2 activity are relatively scarce compared to DIO1. Studies reporting DIO2 inhibition have used human recombinant DIO2 enzyme (Olker et al., 2019), primary human astrocytes (Roberts et al., 2015), rat pituitary (Li et al., 2012), pig liver (Stinckens et al., 2018), Nile tilapia (Oreochromis niloticus) liver (Walpita et al., 2007). Evidence for fish (e.g., zebrafish and fathead minnow) is mostly indirect since DIO enzyme activity is usually not measured in chemical exposure experiments. Houbrechts et al. (2016) showed decreased DIO2 activity in a DIO1-DIO2 knockdown zebrafish at the ages of 3 and 7 days post fertilization together with impaired swim bladder inflation, showing that the enzyme is present, the activity is measurable and impairing its activity has negative effects. Noyes confirmed decreased outer ring deiodination activity in fathead minnows exposed to decabromodiphenyl ether (BDE-209). Walpita et al. (2007) showed decreased DIO2 activity in the liver of Nile tilapia injected with dexamethasone. Stinckens et al. (2018) showed that chemicals with DIO inhibitory potential in pig liver impaired swim bladder inflation in zebrafish, a thyroid hormone regulated process. Six out of seven DIO1 inhibitors impaired posterior chamber inflation, but almost all of these compounds also inhibit DIO2. TCBPA, the only compound that inhibits DIO1 and not DIO2, had no effect on the posterior swim bladder. Based on these results, DIO2 seemed to be more important than DIO1.
In mammals, DIO2 is thought to control the intracellular concentration of T3, while DIO1 is thought to be more important in determining systemic T3 levels. The cells that express DIO2 locally produce T3 that can more rapidly access the thyroid receptors in the nucleus than T3 from plasma (Bianco et al., 2002). For example, DIO2 is highly expressed in the mammalian brain. However, this hypothesis has been challenged. For example, Maia et al. (2005) determined that in a normal physiological situation in humans the contribution of DIO2 to plasma T3 levels is twice that of DIO1. Only in a hyperthyroid state was the contribution of DIO1 higher than that of DIO2. A DIO1 knockout mouse showed normal T3 levels and a normal general phenotype and DIO1 was rather found to play a role in limiting the detrimental effects of conditions that alter normal thyroid function, including hyperthyroidism and iodine deficiency (Schneider et al., 2006). van der Spek et al. concluded that the primary role of DIO1 in vivo is to degrade inactivated TH (van der Spek et al., 2017).
The presence of DIO1 in the liver of teleosts has been a controversial issue and DIO1 function in teleostean and amphibian T3 plasma regulation is unclear (Finnson et al., 1999; Kuiper et al., 2006). In teleosts, DIO2 has a markedly higher activity level compared to other vertebrates and it is expressed in liver (Orozco and Valverde, 2005), suggesting its importance in determining systemic thyroid hormone levels. This could explain why DIO2 inhibition seems to be more important than DIO1 inhibition in determining the adverse outcome in zebrafish (Stinckens et al., 2018).
Life stage: Deiodinase activity is important for all vertebrate life stages. Already during early embryonic development, deiodinase activity is needed to regulate thyroid hormone concentrations and coordinate developmental processes. DIO2 shows more marked changes in expression around the time of the embryo-larval and larval-to-juvenile transition periods during zebrafish development, highlighting its importance for early life stages (Vergauwen et al., 2018).
Sex: This KE is plausibly applicable to both sexes. Deiodinases are important for TH homeostasis and identical in both sexes. Therefore inhibition of deiodinases is not expected to be sex-specific.
Key Event Description
Disruption of the thyroid hormone system is increasingly being recognized as an important toxicity pathway, as it can cause many adverse outcomes. Thyroid hormones do not only play an important role in the adult individual, but they are also critical during embryonic development. Thyroid hormones (THs) play an important role in a wide range of biological processes in vertebrates including growth, development, reproduction, cardiac function, thermoregulation, response to injury, tissue repair and homeostasis. Numerous chemicals are known to disturb thyroid function, for example by inhibiting thyroperoxidase (TPO) or deiodinase (DIO), upregulating excretion pathways or modifying gene expression. The two major thyroid hormones are triiodothyronine (T3) and thyroxine (T4), both iodinated derivatives of tyrosine. Most TH actions depend on the binding of T3 to its nuclear receptors. Active and inactive THs are tightly regulated by enzymes called iodothyronine deiodinases (DIO). The activation occurs via outer ring deiodination (ORD), i.e. removing iodine from the outer, phenolic ring of T4 to form T3, while inactivation occurs via inner ring deiodination (IRD), i.e. removing iodine from the inner tyrosol ring of T4 or T3.
Three types of iodothyronine deiodinases (DIO1-3) have been described in vertebrates that activate or inactivate THs and are therefore important mediators of TH action. All deiodinases are integral membrane proteins of the thioredoxin superfamily that contain selenocysteine in their catalytic centre. Type I deiodinase is capable to convert T4 into T3, as well as to convert reverse T3 (rT3) to 3,3'-Diiodothyronine (3,3’ T2), through outer ring deiodination. rT3, rather than T4, is the preferred substrate for DIO1. furthermore, DIO1 has a very high Km (µM range, compared to nM range for DIO2) (Darras and Van Herck, 2012). Type II deiodinase (DIO2) is only capable of ORD activity with T4 as a preferred substrate (i.e., activation of T4 to T3). DIO3 can inner ring deiodinate T4 and T3 to the inactive forms of THs, rT3 and 3,3’-T2 respectively. DIO2 is a transmembrane protein anchored to the endoplasmic reticulum and the active site faces the perinuclear cytosol. The relative contribution of the DIOs to thyroid hormone levels varies amongst species, developmental stages and tissues.
How it is Measured or Detected
At this time, there are no approved OECD or EPA guideline protocols for measurement of DIO inhibition. Deiodination is the major pathway regulating T3 bioavailability in mammalian tissues. In vitro assays can be used to examine inhibition of deiodinase 2 (DIO2) activity upon exposure to thyroid disrupting compounds.
Several methods for deiodinase activity measurements are available. A first in vitro assay measures deiodinase activities by quantifying the radioactive iodine release from iodine-labelled substrates, depending on the preferred substrates of the isoforms of deiodinases (Forhead et al., 2006; Pavelka, 2010; Houbrechts et al., 2016; Stinckens et al., 2018). Each of these assays requires a source of deiodinase which can be obtained for example using unexposed pig liver tissue (available from slaughterhouses) or rat liver tissue. Olker et al. (2019) on the other hand used an adenovirus expression system to produce the DIO2 enzyme and developed an assay for nonradioactive measurement of iodide released using the Sandell-Kolthoff method, a photometric method based on Ce4+ reduction (Renko et al., 2012). This assay was then used to screen the ToxCast Phase 1 chemical library. The specific synthesis of DIO2 through the adenovirus expression system provides an important advantage over other methods where activity of the different deiodinase isoforms needs to be distinguished in other ways, such as based on differences in enzyme kinetics.
Measurements of in vivo deiodinase activity in tissues collected from animal experiments are scarce. Noyes et al. (2011) showed decreased rate of outer ring deiodination (mediated by DIO1 and DIO2) in whole fish microsomes after exposure to BDE-209. After incubation with the substrate, thyroid hormone levels were measured using LC-MS/MS. Houbrechts et al. (2016) confirmed DIO2 deiodination activity in a DIO1-DIO2 knockdown zebrafish at the ages of 3 and 7 days post fertilization. Decreased T3 levels are often used as evidence of DIO inhibition, for example after exposure to iopanoic acid, in fish species such as zebrafish (Stinckens et al., 2020) and fathead minnow (Cavallin et al., 2017). It should be noted that it is difficult to make the distinction between decreased T3 levels caused by outer ring deiodination mediated by DIO2 inhibition or DIO1 inhibition.
References
Bianco, A.C., Salvatore, D., Gereben, B., Berry, M.J., Larsen, P.R., 2002. Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeiodinases. Endocrine Reviews 23, 38-89.
Cavallin JE, Ankley GT, Blackwell BR, Blanksma CA, Fay KA, Jensen KM, Kahl MD, Knapen D, Kosian PA, Poole ST et al. 2017. Impaired swim bladder inflation in early life stage fathead minnows exposed to a deiodinase inhibitor, iopanoic acid. Environmental Toxicology and Chemistry. 36(11):2942-2952.
Darras, V.M., Van Herck, S.L.J., 2012. Iodothyronine deiodinase structure and function: from ascidians to humans. Journal of Endocrinology 215, 189-206.
Forhead, A.J., Curtis, K., Kaptein, E., Visser, T.J., Fowden, A.L., 2006. Developmental control of iodothyronine deiodinases by cortisol in the ovine fetus and placenta near term. Endocrinology 147, 5988-5994.
Houbrechts, A.M., Delarue, J., Gabriels, I.J., Sourbron, J., Darras, V.M., 2016. Permanent Deiodinase Type 2 Deficiency Strongly Perturbs Zebrafish Development, Growth, and Fertility. Endocrinology 157, 3668-3681.
Li, N.N., Jiang, Y.Q., Shan, Z.Y., Teng, W.P., 2012. Prolonged high iodine intake is associated with inhibition of type 2 deiodinase activity in pituitary and elevation of serum thyrotropin levels. British Journal of Nutrition 107, 674-682.
Noyes PD, Hinton DE, Stapleton HM. 2011. Accumulation and debromination of decabromodiphenyl ether (bde-209) in juvenile fathead minnows (pimephales promelas) induces thyroid disruption and liver alterations. Toxicological Sciences. 122(2):265-274.
Olker, J.H., Korte, J.J., Denny, J.S., Hartig, P.C., Cardon, M.C., Knutsen, C.N., Kent, P.M., Christensen, J.P., Degitz, S.J., Hornung, M.W., 2019. Screening the ToxCast Phase 1, Phase 2, and e1k Chemical Libraries for Inhibitors of Iodothyronine Deiodinases. Toxicological Sciences 168, 430-442.
Orozco, A., Valverde, R.C., 2005. Thyroid hormone deiodination in fish. Thyroid 15, 799-813.
Pavelka, S., 2010. Radiometric enzyme assays: development of methods for extremely sensitive determination of types 1, 2 and 3 iodothyronine deiodinase enzyme activities. Journal of Radioanalytical and Nuclear Chemistry 286, 861-865.
Renko, K., Hoefig, C.S., Hiller, F., Schomburg, L., Kohrle, J., 2012. Identification of Iopanoic Acid as Substrate of Type 1 Deiodinase by a Novel Nonradioactive Iodide-Release Assay. Endocrinology 153, 2506-2513.
Renko, K., Schache, S., Hoefig, C.S., Welsink, T., Schwiebert, C., Braun, D., Becker, N.P., Kohrle, J., Schomburg, L., 2015. An Improved Nonradioactive Screening Method Identifies Genistein and Xanthohumol as Potent Inhibitors of Iodothyronine Deiodinases. Thyroid 25, 962-968.
Roberts, S.C., Bianco, A.C., Stapleton, H.M., 2015. Disruption of Type 2 Iodothyronine Deiodinase Activity in Cultured Human Glial Cells by Polybrominated Diphenyl Ethers. Chemical Research in Toxicology 28, 1265-1274.
Schneider, M.J., Fiering, S.N., Thai, B., Wu, S.Y., St Germain, E., Parlow, A.F., St Germain, D.L., Galton, V.A., 2006. Targeted disruption of the type 1 selenodeiodinase gene (Dio1) results in marked changes in thyroid hormone economy in mice. Endocrinology 147, 580-589.
Stinckens, E., Vergauwen, L., Ankley, G.T., Blust, R., Darras, V.M., Villeneuve, D.L., Witters, H., Volz, D.C., Knapen, D., 2018. An AOP-based alternative testing strategy to predict the impact of thyroid hormone disruption on swim bladder inflation in zebrafish. Aquatic Toxicology 200, 1-12.
Stinckens E, Vergauwen L, Blackwell BR, Anldey GT, Villeneuve DL, Knapen D. 2020. Effect of thyroperoxidase and deiodinase inhibition on anterior swim bladder inflation in the zebrafish. Environmental Science & Technology. 54(10):6213-6223.
van der Spek, A.H., Fliers, E., Boelen, A., 2017. The classic pathways of thyroid hormone metabolism. Molecular and Cellular Endocrinology 458, 29-38.
Vergauwen, L., Cavallin, J.E., Ankley, G.T., Bars, C., Gabriels, I.J., Michiels, E.D.G., Fitzpatrick, K.R., Periz-Stanacev, J., Randolph, E.C., Robinson, S.L., Saari, T.W., Schroeder, A.L., Stinckens, E., Swintek, J., Van Cruchten, S.J., Verbueken, E., Villeneuve, D.L., Knapen, D., 2018. Gene transcription ontogeny of hypothalamic-pituitary-thyroid axis development in early-life stage fathead minnow and zebrafish. General and Comparative Endocrinology 266, 87-100.
Walpita, C.N., Grommen, S.V., Darras, V.M., Van der Geyten, S., 2007. The influence of stress on thyroid hormone production and peripheral deiodination in the Nile tilapia (Oreochromis niloticus). Gen Comp Endocrinol 150, 18-25.
Event: 1656: Antagonism, Thyroid Receptor
Short Name: TR Antagnoism
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Molecular |
Domain of Applicability
Taxonomic Applicability Life Stage Applicability| Life Stage | Evidence |
|---|---|
| During development and at adulthood | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
Key Event Description
Thyroid hormone receptors (TR) are a nuclear receptors that are activitated by binding of the thyroid hormones triiodothyronine (T3) and thyroxine (T4). The majority of TH bound to TR being T3 due to its 10-fold higher affinity. Bound receptors, homodimerized or heterodimerized with retinoic acid, bind to thyroid response elements and regulate gene expression by either increasing or decreasing tragent gene transcription activity. Important to note is ligand free TR can form complexes with corepressors to inhibit gene expression. There are two major thyroid hormone receptor subtypes, thyroid receptor alpha(TRα) and thyroid receptor beta (TRβ). There are two subtypes for each, TRb1, TRb2, TRa1, and TRa2. Notably, the carboxy-terminal structure of TRalpha2 prevents hormone binding and transscription (Sinha and Yen, 2018). There are a large number of genes regualated by TH. These include genes involved in
Both TRa and TRb are known to be expressed during neurodevelopment (ref).
The predominate TR form during brain develop is TRa1expression of the
Sinha R, Yen PM. Cellular Action of Thyroid Hormone. [Updated 2018 Jun 20]. In: Feingold KR, Anawalt B, Boyce A, et al., editors. Endotext [Internet]. South Dartmouth (MA): MDText.com, Inc.; 2000-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK285568/
List of Key Events in the AOP
Event: 2093: Brain thyroid hormone (T4/T3) decreased
Short Name: Decreased brain T4/T3
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Organ |
Event: 381: Reduced levels of BDNF
Short Name: BDNF, Reduced
Key Event Component
| Process | Object | Action |
|---|---|---|
| gene expression | brain-derived neurotrophic factor | decreased |
| secretion | brain-derived neurotrophic factor | decreased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Molecular |
Cell term
| Cell term |
|---|
| neural cell |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| human | Homo sapiens | High | NCBI |
| rat | Rattus norvegicus | High | NCBI |
| mouse | Mus musculus | High | NCBI |
| Life Stage | Evidence |
|---|---|
| During brain development | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
BDNF plays a critical role in normal brain development in most vertebrates, primarily documented empirically in mammalian species. Klein et al. (2011) examined blood, serum, plasma and brain-tissue and measured BDNF levels in three different mammalian species: rat, pig, and mouse, using an ELISA method (Aid et al., 2007), whereas Trajkovska et al. 2007 determined BDNF levels in human blood.
There is compelling data that demonstrates the role of BDNF in brain development for many other taxa, including fish where it acts as neurotrophic factor in controlling cell proliferation (D'Angelo L et al., 2014; Heinrich and Pagtakhan, 2004) and birds where BDNF influences development of the brain area that involved in the song control (Brenowitz 2013) and the addition of new neurons to a cortical nucleus in adults . In the Xenopus visual system, BDNF acts as neurotrophic factor that mediates synaptic differentiation and maturation of the retinotectal circuit through cell autonomous TrkB signaling on retinal ganglion cells (Sanchez et al., 2006; Marshak et al., 2007).
Key Event Description
BDNF (Brain-derived neurotrophic factor) plays a critical role in normal brain development in most vertebrates, primarily documented empirically in mammalian species. Klein et al. (2011) examined blood, serum, plasma and brain-tissue and measured BDNF levels in three different mammalian species: rat, pig, and mouse, using an ELISA method (Aid et al., 2007), whereas Trajkovska et al. 2007 determined BDNF levels in human blood.
There is compelling data that demonstrates the role of BDNF in brain development for many other taxa, including fish where it acts as neurotrophic factor in controlling cell proliferation (D'Angelo L et al., 2014; Heinrich and Pagtakhan, 2004) and birds where BDNF influences development of the brain area that involved in the song control (Brenowitz 2013) and the addition of new neurons to a cortical nucleus in adults . In the Xenopus visual system, BDNF acts as neurotrophic factor that mediates synaptic differentiation and maturation of the retinotectal circuit through cell autonomous tropomycin receptor kinase B also known as tyrosine receptor kinase B (TrkB) signaling on retinal ganglion cells (Sanchez et al., 2006; Marshak et al., 2007).
Biological state: BDNF belongs to a family of closely related neurotrophic factors named neurotrophins and is widely expressed in the developing and mature central nervous system (CNS). In the rodent cortex, postnatal BDNF expression is initially low but slowly increases to reach high levels around weaning. Therefore, BDNF expression peaks at a time when both structural and functional maturation of cortical circuitry occurs. During postnatal development, BDNF levels are dynamically regulated, in part by neuronal activity dependent mechanisms (Waterhouse and Xu, 2009). Glutamate has been shown to increase the transcription and release of BDNF. Indeed, BDNF is synthesized, stored and released from glutamatergic neurons (Lessmann et al., 2003).
Biological compartments: BDNF initially is synthesized as precursor proteins (proBDNF), which is processed intracellularly to be transformed in its mature form (mBDNF) after proteolytically cleaved in the synaptic cleft by plasmin which is a protease activated by tissue plasminogen activator (tPA) (Cohen-Cory et al., 2010). proBDNF is constantly secreted while tPA release and mBDNF production depends on neuronal excitation (Head et al., 2009). Storage and activity-dependent release of BDNF has been demonstrated in both dendrites and axon terminals (Waterhouse and Xu, 2009). More specifically, in hippocampus, BDNF appears to be stored in dendritic processes of neurons (Balkowiec and Katz, 2002). BDNF is abundant in cerebellum and cortex and has also been measured in cerebrospinal fluid (CSF) (Zhang et al., 2008), whole blood, plasma, serum (plasma without clotting factors) and platelets (Trajkovska et al., 2007). BDNF has been found to be produced by astrocytes under both physiological and pathological conditions (Endo, 2005; Coco et al., 2013; Nelson and Alkon, 2014).
In humans (Pruunsild et al., 2007), mBDNF is sequestered in platelets, consequently BDNF can reach all tissues and organs. Lymphocytic cells have been shown to express BDNF in vitro similarly to eosinophils, dendritic cells, and endothelial cells. The visceral and airway epithelium are also significant sources of BDNF. Female reproductive system including ovaries, placenta and uterus also express BDNF (Wessels et al., 2014).
General role in biology: The biological functions of mBDNF are mediated by binding to tyrosine kinase B (TrkB) receptor that leads to the activation of three major intracellular signalling pathways, including MAPK, PI3K and PLCγ1 (Soulé et al., 2006). TrkB-mediated signaling regulates gene transcription in the nucleus through the activation of several transcription factors. These genes are involved in neurite outgrowth, synaptogenesis, synapse maturation and stabilization (Pang et al., 2004; Lu et al., 2005; Nelson and Alkon, 2014).
On the other hand, proBDNF binds to the p75 neurotrophin receptor (p75NTR) and activates RhoA, a small GTPase that regulates actin cytoskeleton polymerization leading to inhibition of axonal elongation, growth cone collapse, and apoptosis (Dubreuil et al., 2003; Yamauchi et al., 2004; Head et al., 2009).
How it is Measured or Detected
Methods that have been previously reviewed and approved by a recognized authority should be included in the Overview section above. All other methods, including those well established in the published literature, should be described here. Consider the following criteria when describing each method: 1. Is the assay fit for purpose? 2. Is the assay directly or indirectly (i.e. a surrogate) related to a key event relevant to the final adverse effect in question? 3. Is the assay repeatable? 4. Is the assay reproducible?
No OECD methods are available to measure BDNF protein and mRNA levels. Measuring BDNF levels changes in the brain, especially when low, at the boarder to be significant are technically difficult. Depending on the tissue or fluid measurements distinct methods are used.
Brain tissue: BDNF protein levels can be measured by commercial available antibody sandwich ELISA kits, Western blotting, immunohistochemistry and immunofluorescence. BDNF primers for different exons are available to determine mRNA levels by RT-PCR. The Bdnf gene consists of multiple alternative exons (ten in human, eight in rodents and six in lower vertebrates), and a single exon coding for the entire pro-BDNF protein (Cohen-Cory et al., 2010).
Cerebro-spinal fluid (CSF): There are available commercial antibody sandwich ELISA kits (Trajkovska et al., 2007) and immunobead-based multiplex assays for high throughput screening (Zhang et al., 2008).
Whole blood, serum, plasma and platelets: There are several commercial double antibody sandwich ELISA kits that can be used for identification of BDNF levels in biological fluids (Trajkovska et al., 2007).
Methodological considerations that have to be taken into account during sample preparation and measurement of BDNF by ELISA have been recently reviewed in Elfving et al. 2010. A study measuring BDNF by a commercially available ELISA kit in various tissues and biological liquids derived from distinct species revealed that BDNF is undetectable in mouse blood and pig plasma (Klein et al., 2011). This study also showed that in most cases BDNF levels are comparable to levels reported in humans and that there is positive correlation between blood BDNF levels and hippocampal BDNF levels in rats and pigs (Klein et al., 2011).
References
Aid T, Kazantseva A, Piirsoo M, Palm K, Timmusk T. (2007) Mouse and rat BDNF gene structure and expression revisited. J Neurosci Res. 85: 525-535.
Balkowiec A, Katz DM. (2002) Cellular mechanisms regulating activity-dependent release of native brain-derived neurotrophic factor from hippocampal neurons. J Neurosci. 22: 10399-10407.
Brenowitz EA. (2013) Testosterone and brain-derived neurotrophic factor interactions in the avian song control system. Neuroscience 239: 115-123.
Coco M, Caggia S, Musumeci G, Perciavalle V, Graziano AC, Pannuzzo G, Cardile V. (2013) Sodium L-lactate differently affects brain-derived neurothrophic factor, inducible nitric oxide synthase, and heat shock protein 70 kDa production in human astrocytes and SH-SY5Y cultures.J Neurosci Res. 91: 313-320.
Cohen-Cory S, Kidane AH, Shirkey NJ, Marshak S. (2010) Brain-derived neurotrophic factor and the development of structural neuronal connectivity. Dev Neurobiol. 70: 271-288.
D'Angelo L, De Girolamo P, Lucini C, Terzibasi ET, Baumgart M, Castaldo L, Cellerino A (2014). Brain-derived neurotrophic factor: mRNA expression and protein distribution in the brain of the teleost Nothobranchius furzeri. J Comp Neurol. 1;522(5):1004-30.
Dubreuil CI, Winton MJ, McKerracher L. (2003) Rho activation patterns after spinal cord injury and the role of activated Rho in apoptosis in the central nervous system. J Cell Biol. 162: 233-243.
Elfving B, Plougmann PH, Wegener G. (2010) Detection of brain-derived neurotrophic factor (BDNF) in rat blood and brain preparations using ELISA: pitfalls and solutions. J Neurosci Methods 187: 73-77.
Endo T. (2005) Glycans and glycan-binding proteins in brain: galectin-1-induced expression of neurotrophic factors in astrocytes. Curr Drug Targets. 6:427-436.
Head BP, Patel HH, Niesman IR, Drummond JC, Roth DM, Patel PM. (2009) Inhibition of p75 neurotrophin receptor attenuates isoflurane-mediated neuronal apoptosis in the neonatal central nervous system. Anesthesiology 110: 813-825.
Heinrich G, Pagtakhan CJ. (2004) Both 5' and 3' flanks regulate Zebrafish brain-derived neurotrophic factor gene expression. BMC Neurosci. 5: 19.
Klein AB, Williamson R, Santini MA, Clemmensen C, Ettrup A, Rios M, Knudsen GM, Aznar S. (2011) Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int J Neuropsychopharmacol. 14: 347-353.
Lessmann V, Gottmann K, Malcangio M. (2003) Neurotrophin secretion: current facts and future prospects. Prog Neurobiol. 69: 341-374.
Lu B, Pang PT, Woo NH. (2005) The yin and yang of neurotrophin action. Nat Rev Neurosci. 6: 603-614.
Marshak S, Nikolakopoulou AM, Dirks R, Martens GJ, Cohen-Cory S (2007)Cell-autonomous TrkB signaling in presynaptic retinal ganglion cells mediates axon arbor growth and synapse maturation during the establishment of retinotectal synaptic connectivity. J Neurosci 27:2444 –2456.
Nelson TJ, Alkon DL. (2014) Molecular regulation of synaptogenesis during associative learning and memory. Brain Res. pii: S0006-8993(14)01660-6. doi: 10.1016/j.brainres.2014.11.054.
Pang PT, Teng HK, Zaitsev E, Woo NT, Sakata K, Zhen S, Teng KK, Yung WH, Hempstead BL, Lu B. (2004) Cleavage of proBDNF by tPA/plasmin is essential for long-term hippocampal plasticity. Science. 306: 487-491.
Pruunsild P, Kazantseva A, Aid T, Palm K, Timmusk T. (2007) Dissecting the human BDNF locus: bidirectional transcription, complex splicing, and multiple promoters. Genomics. 90: 397-406.
Sanchez AL, Matthews BJ, Meynard MM, Hu B, Javed S, Cohen Cory S (2006) BDNF increases synapse density in dendrites of developing tectal neurons in vivo. Development 133:2477–2486.
Soulé J, Messaoudi E, Bramham CR. (2006) Brain-derived neurotrophic factor and control of synaptic consolidation in the adult brain. Biochem Soc Trans. 34 :600-604.
Trajkovska V, Marcussen AB, Vinberg M, Hartvig P, Aznar S, Knudsen GM. (2007) Measurements of brain-derived neurotrophic factor: methodological aspects and demographical data. Brain Res Bull. 73: 143-149.
Waterhouse EG, Xu B. (2009) New insights into the role of brain-derived neurotrophic factor in synaptic plasticity. Mol Cell Neurosci. 42: 81-89.
Wessels JM, Wu L, Leyland NA, Wang H, Foster WG. (2014) The Brain-Uterus Connection: Brain Derived Neurotrophic Factor (BDNF) and Its Receptor (Ntrk2) Are Conserved in the Mammalian Uterus. PLoS ONE 9: e94036.
Yamauchi J, Chan JR, Shooter EM. (2004) Neurotrophins regulate Schwann cell migration by activating divergent signaling pathways dependent on Rho GTPases. Proc Natl Acad Sci U S A. 101: 8774-8779.
Zhang J, Sokal I, Peskind ER, Quinn JF, Jankovic J, Kenney C, Chung KA, Millard SP, Nutt JG, Montine TJ. (2008) CSF multianalyte profile distinguishes Alzheimer and Parkinson diseases. Am J Clin Pathol. 129: 526-529.
Event: 385: Decrease of synaptogenesis
Short Name: Synaptogenesis, Decreased
Key Event Component
| Process | Object | Action |
|---|---|---|
| synapse assembly | synapse | decreased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Cellular |
Cell term
| Cell term |
|---|
| neuron |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| human | Homo sapiens | High | NCBI |
| rat | Rattus norvegicus | High | NCBI |
| mouse | Mus musculus | High | NCBI |
| Life Stage | Evidence |
|---|---|
| During brain development | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
The mechanisms governing synapse formation is considered conserved among both vertebrates and invertebrates (Munno and Syed, 2003). Invertebrates have served as simple animal models to study synapse formation. Indeed, Colón-Ramos (2009) has recently reviewed the early developmental events that take place in the process of synaptogenesis pointing out the importance of this process in neural network formation and function. The experimental evaluation of synaptogenesis has been performed using invertebrates and in particular C. elegans and Drosophila as well as vertebrates (Colón-Ramos, 2009).
This vulnerable period of synaptogenesis appears to happen in different developmental stages across species. For example, in rodents primarily synaptogenesis occurs during the first two weeks after birth (Bai et al., 2013). For rhesus monkeys, this period ranges from approximately 115-day gestation up to PND 60 (Bai et al., 2013). In humans, it starts from the third trimester of pregnancy and continues 2-3 years following birth (Bai et al., 2013).
Key Event Description
Biological state: Synaptogenesis is a multi-step process that is crucial for brain development and involves the formation of synapses. It follows axonal migration, at which stage presynaptic and postsynaptic differentiation occurs (Garner et al., 2002). "Synaptic assembly" that refers to the gathering of the appropriate components and "synaptic formation" that is defined by the mechanisms involved in recruitment of molecules required for differentiation, stabilization and maturation of synapse, are the main phases that characterise synaptogenesis (Colón-Ramos, 2009). Elimination is a physiological step involved in synaptogenesis regarding the synapses that fail to get stabilised and mature.
The first step is the recognition and the establishment of contact between an axon and a dendritic spine in which pre- and postsynaptic neurons play important role. The presynaptic differentiation occurs followed by excretion of neurotransmitters that bind to appropriate receptors located on the target spine. However, a postsynaptic neuron does not passively receive guidance from a presynaptic axon but are the same dendritic filopodia that gradually are transformed into spines that select and engage their presynaptic neurons. The transformation of dendritic filopodia into dendritic spines that involves the expression of the whole postsynaptic machinery such as postsynaptic density (PSD), receptor subunits, scaffolding proteins and actin cytoskeleton, is the first step to give nascent synapses. However, to become functional and mature these synapses need an important number of cell-cell interactions, including stimulation from glutamatergic synapses as well as the influence of neurotrophic factors (Munno and Syed, 2003).
However, all this is true for glutamatergic synapses because GABAergic synapses do not appear in dendritic spines, but rather form on dendritic shafts, nerve cell somata and axon initial segments. These inhibitory synapses besides their distinct location are also structurally different compared to excitatory synapses (reviewed in Gatto and Broadie, 2010).
Biological compartments: Synaptogenesis is spatially and temporally strictly controlled process. It does not happen in a uniform way in all brain regions and there important differences between the times of appearance of the main two types of synapses (reviewed in Erecinska et al., 2004). For example, in rat hippocampus excitatory synapses are well established or fully mature within the two first postnatal weeks, whereas inhibitory synapses cannot be found prior to PND 18, after which it increases steadily to reach adult levels at PND 28. In addition, in rat neostriatal neurons the excitatory responses to both cortical and thalamic stimuli can be observed by PND 6, but the long-lasting hyperpolarization and late depolarization is never seen before PND 12.
Structural remodelling of synapses and formation of new synaptic contacts has been postulated as a possible mechanism underlying the late phase of long-term potentiation (LTP), a form of plasticity which is involved in learning and memory. LTP induction results in a sequence of morphological changes consisting of a transient remodelling of the postsynaptic membrane followed by a marked increase in the proportion of axon terminals contacting two or more dendritic spines. Three-dimensional reconstruction revealed that these spines arose from the same dendrite. As pharmacological blockade of LTP prevented these morphological changes, it is suggested that LTP is associated with the formation of new, mature and probably functional synapses contacting the same presynaptic terminal and thereby duplicating activated synapses (Erik et al., 2006).
In human, synaptogenesis does not happen at the same time in all brain regions, as the prefrontal cortex lags behind in terms of synapse formation compared to the auditory and visual cortices. In contrast, synaptogenesis appears to proceed concurrently in different brain areas for rhesus monkey (Erecinska et al., 2004).
General role in biology: The period of rapid synaptogenesis or the so-called brain growth spurt is considered one of the most important processes that take place during brain development (Garner et al., 2002). This process is crucial not only in neurodevelopment but also plays a vital role in synaptic plasticity, learning and memory and adaptation throughout life. Without this process no complex brain network can be established as synapse is the fundamental unit of connectivity and communication between neurons (Tau and Peterson, 2010). Cell adhesion represents the most direct way of coordinating synaptic connectivity in the brain. Recent evidence highlights the importance of a trans-synaptic interaction between postsynaptic neuroligins and presynaptic neurexins. These transmembrane molecules bind each other extracellularly to promote adhesion between dendrites and axons, facilitating synapse establishment (Dean and Dresbach, 2006). Furthermore, the number of excitatory versus inhibitory synapses created at single neuron dictates neuronal excitability and function (Schummers et al., 2002).
How it is Measured or Detected
Methods that have been previously reviewed and approved by a recognized authority should be included in the Overview section above. All other methods, including those well established in the published literature, should be described here. Consider the following criteria when describing each method: 1. Is the assay fit for purpose? 2. Is the assay directly or indirectly (i.e. a surrogate) related to a key event relevant to the final adverse effect in question? 3. Is the assay repeatable? 4. Is the assay reproducible?
There is no OECD advised method for measuring synaptogenesis.
Anatomical methods can be used to structurally estimate the number of excitatory or inhibitory synapses. Immunostaining can be employed with specific antibodies that recognize vesicular glutamate transporters (VGLUTs) and the postsynaptic density protein-95 kDa (PSD-95) that are characteristic of excitatory synapses, while inhibitory synapses are identified by the presence of the vesicular GABA (VGAT) and vesicular inhibitory amino acid (VIAAT) transporters and the postsynaptic adaptor protein gephryin (Gatto and Broadie, 2010). There are commercial available synaptogenesis assay kits that rely on the immunostaining of cells with MAP-2, PSD-95 and synaptophysin. Some other presynaptic (Bassoon) and postsynaptic (ProSAP1/Shank2) markers have been suggested and showed to correlate well with the ultrastructural studies in cultured hippocampus primary cells (Grabrucker et al., 2009). Electron microscopy can also be applied to assess the prevalence of excitatory and inhibitory synapses amongst convergent contacts (Megias et al., 2001). Recently, a high content image analysis based on RNAi screening protocols has been suggested as a useful tool to create imaging algorithm for use in both in vitro and in vivo synaptic punctae analysis (Nieland et al., 2014).
References
Bai X, Twaroski D, Bosnjak ZJ. (2013) Modeling anesthetic developmental neurotoxicity using human stem cells. Semin Cardiothorac Vasc Anesth. 17: 276-287.
Colón -Ramos DA. (2009) Synapse formation in developing neural circuits. Curr Top Devel Biol. 87: 53-79.
Dean C, Dresbach T. (2006) Neuroligins and neurexins: linking cell adhesion, synapse formation and cognitive function. Trends Neurosci. 29:21-29.
Erecinska M, Cherian S, Silver IA. (2004) Energy metabolism in mammalian brain during development. Prog Neurobiol. 73: 397-445.
Erik I. Charyc, Barbara F. Akum, Joshua S. Goldber, Rebecka J. Jörnsten, Christopher Rongo, James Q. Zheng and Bonnie L. Firestein. Activity-Independent Regulation of Dendrite Patterning by Postsynaptic Density Protein PSD-95. Journal of Neuroscience 2006, 26(40): 10164-10176.
Garner CC, Zhai RC, Gundelfinger ED, Ziv NE. (2002) Molecular mechanisms of CNS synaptogenesis. Cell Press 25: 243-250.
Gatto CL, Broadie K. (2010) Genetic controls balancing excitatory and inhibitory synaptogenesis in neurodevelopmental disorder models. Front Syn Neurosci. 2: 4.
Grabrucker A, Vaida B, Bockmann J, Boeckers TM. (2009) Synaptogenesis of hippocampal neurons in primary cell culture. Cell Tissue Res. 338: 333-341.
Megias M, Emri Z, Freund TF, Gulyas AI. (2001) Total number and distribution of inhibitory and excitatory synapses on hippocampal CA1 pyramidal cells. Neuroscience 102: 527-540.
Munno DW, Syed NI. (2003) Synaptogenesis in the CNS: an odyssey from wiring together to firing together. J Physiol. 552: 1-11.
Nieland TJF, Logan DJ, Saulnier J, Lam D, Johnson C, et al. (2014) High Content Image Analysis Identifies Novel Regulators of Synaptogenesis in a High-Throughput RNAi Screen of Primary Neurons. PLoS ONE. 9: e91744.
Schummers J, Mariño J, Sur M. (2002) Synaptic integration by V1 neurons depends on location within the orientation map. Neuron. 36: 969-978.
Tau GZ, Peterson BS. (2010) Normal Development of Brain Circuits. Neuropsychopharmacology 35: 147-168.
Event: 1505: Cell cycle, disrupted
Short Name: Cell cycle, disrupted
Key Event Component
| Process | Object | Action |
|---|---|---|
| regulation of cell cycle | cell cycle-related cyclin | disrupted |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Cellular |
Cell term
| Cell term |
|---|
| cell |
Organ term
| Organ term |
|---|
| organ |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | High | NCBI |
| Mus musculus | Mus musculus | High | NCBI |
| Life Stage | Evidence |
|---|---|
| Not Otherwise Specified | Moderate |
| Sex | Evidence |
|---|---|
| Unspecific | High |
The histone gene expression alters in each phase of the cell cycle in human HeLa cells (Homo sapiens) [Heintz et al., 1982].
Key Event Description
The disruption of the cell cycle leads to a decrease in cell number. The cell cycle consists of G1, S, G2, M, and G0 phases. The cell cycle regulation is disrupted by the cell cycle arrest in certain cell cycle phases. The histone gene expression is regulated in cell cycle phases [Heintz et al., 1983].
How it is Measured or Detected
The percentage of cells at G1, G0, S, and G2/M phases can be detected by flow cytometry [Li et al., 2013]. Cell cycle distribution was analyzed by fluorescence-activated cell sorter (FACS) analysis with a Partec PAS-II sorter [Zupkovitz et al., 2010]. The four cell-cycle phases in living cells can be measured with four-color fluorescent proteins using live-cell imaging [Bajar et al., 2016]. The incorporation of [3H]deoxycytidine or [3H]thymidine into cell DNA during the S phase can be monitored as DNA synthesis [Heintz et al., 1982].
References
Bajar, B.T. et al. (2016), "Fluorescent indicators for simultaneous reporting of all four cell cycle phases", Nat Methods 13:993-996
Heintz, N. et al. (1983), "Regulation of human histone gene expression: Kinetics of accumulation and changes in the rate of synthesis and in the half-lives of individual histone mRNAs during the HeLa cell cycle", Molecular and Cellular Biology 3:539-550
Li, Q. et al. (2013), "Glyphosate and AMPA inhibit cancer cell growth through inhibiting intracellular glycine synthesis", Drug Des Devel Ther 7:635-643
Event: 2105: Decreased, myelin basic protein expression in oligodendrocytes
Short Name: Decreased, mbp expression in oligodendrocytes
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Molecular |
Event: 2266: Demyelination, increased
Short Name: Demyelination, increased
Key Event Component
| Process | Object | Action |
|---|---|---|
| demyelination | increased | |
| CNS demyelination | increased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Cellular |
Organ term
| Organ term |
|---|
| nervous system |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | NCBI |
| Sex | Evidence |
|---|---|
| Unspecific |
Key Event Description
Demyelination is defined by the loss of myelin sheaths in nervous tissue, typically following insult from injury or disease. Demyelination is initiated by fractioning of myelin lamellae followed by removal of the fragments by proteolytic and lipolytic enzymes that can digest the myelin pieces (Cuzner & Norton, 1996; Höftberger & Lassmann, 2017). Considering myelin functions to maintain axon functionality and survival, once myelin is lost neurodegeneration ensues (Ohno & Ikenaka, 2019). Demyelinating lesions can occur anywhere within the CNS including on myelin surrounding axons of both sensory and motor neurons (Höftberger & Lassmann, 2017).
References
Cuzner, M. L., & Norton, W. T. (1996). Biochemistry of Demyelination. Brain Pathology, 6(3), 231-242.
Höftberger, R., & Lassmann, H. (2017). Inflammatory demyelinating diseases of the central nervous system. Handbook of Clinical Neurology, 145, 263–283.
Ohno, N., & Ikenaka, K. (2019). Axonal and neuronal degeneration in myelin diseases. Neuroscience Research, 139, 48-57.
Event: 2378: Impaired axonal insulation
Short Name: Impaired axonal insulation
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:610 - Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | KeyEvent |
Biological Context
| Level of Biological Organization |
|---|
| Tissue |
Event: 2377: Decrease, neural network maturation
Short Name: neural network formation
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:610 - Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | KeyEvent |
Biological Context
| Level of Biological Organization |
|---|
| Tissue |
Event: 386: Decrease of neuronal network function
Short Name: Neuronal network function, Decreased
Key Event Component
| Process | Object | Action |
|---|---|---|
| synaptic signaling | decreased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Organ |
Organ term
| Organ term |
|---|
| brain |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| humans | Homo sapiens | High | NCBI |
| rat | Rattus norvegicus | High | NCBI |
| mice | Mus sp. | High | NCBI |
| cat | Felis catus | High | NCBI |
| Life Stage | Evidence |
|---|---|
| During brain development | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
In vitro studies in brain slices applying electrophysiological techniques showed significant variability among species (immature rats, rabbits and kittens) related to synaptic latency, duration, amplitude and efficacy in spike initiation (reviewed in Erecinska et al., 2004).
Key Event Description
Biological state: There are striking differences in neuronal network formation and function among the developing and mature brain. The developing brain shows a slow maturation and a transient passage from spontaneous, long-duration action potentials to synaptically-triggered, short-duration action potentials.
Furthermore, at this precise developmental stage the neuronal network is characterised by "hyperexcitability”, which is related to the increased number of local circuit recurrent excitatory synapses and the lack of γ-amino-butyric acid A (GABAA)-mediated inhibitory function that appears much later. This “hyperexcitability” disappears with maturation when pairing of the pre- and postsynaptic partners occurs and synapses are formed generating population of postsynaptic potentials and population of spikes followed by developmental GABA switch. Glutamatergic neurotransmission is dominant at early stages of development and NMDA receptor-mediated synaptic currents are far more times longer than those in maturation, allowing more calcium to enter the neurons. The processes that are involved in increased calcium influx and the subsequent intracellular events seem to play a critical role in establishment of wiring of neural circuits and strengthening of synaptic connections during development (reviewed in Erecinska et al., 2004). Neurons that do not receive glutaminergic stimulation are undergoing developmental apoptosis.
During the neonatal period, the brain is subject to profound alterations in neuronal circuitry due to high levels of synaptogenesis and gliogenesis. For example, in neuroendocrine regions such as the preoptic area-anterior hypothalamus (POA-AH), the site of gonadotropin-releasing hormone (GnRH) system is developmentally regulated by glutamatergic neurons. The changes in the expression of the N-methyl-D-aspartate (NMDA) receptor subunits NR1 and NR2B system begin early in postnatal development, before the onset of puberty, thereby playing a role in establishing the appropriate environment for the subsequent maturation of GnRH neurons (Adams et al., 1999).
Biological compartments: Neural network formation and function happen in all brain regions but it appears to onset at different time points of development (reviewed in Erecinska et al., 2004). Glutamatergic neurotransmission in hippocampus is poorly developed at birth. Initially, NMDA receptors play important role but the vast majority of these premature glutamatergic synapses are “silent” possibly due to delayed development of hippocampal AMPA receptors. In contrast, in the cerebral cortex the maturation of excitatory glutamatergic neurotransmission happens much earlier. The “silent” synapses disappear by PND 7-8 in both brain regions mentioned above.
There is strong evidence suggesting that NMDA receptor subunit composition controls synaptogenesis and synapse stabilization (Gambrill and Barria, 2011). It is established fact that during early postnatal development in the rat hippocampus, synaptogenesis occurs in parallel with a developmental switch in the subunit composition of NMDA receptors from NR2B to NR2A. It is suggested that early expression of NR2A in organotypic hippocampal slices reduces the number of synapses and the volume and dynamics of spines. In contrast, overexpression of NR2B does not affect the normal number and growth of synapses. However, it does increase spine motility, adding and retracting spines at a higher rate. The C terminus of NR2B, and specifically its ability to bind CaMKII, is sufficient to allow proper synapse formation and maturation. Conversely, the C terminus of NR2A was sufficient to stop the development of synapse number and spine growth. These results indicate that the ratio of synaptic NR2B over NR2A controls spine motility and synaptogenesis, and suggest a structural role for the intracellular C terminus of NR2 in recruiting the signalling and scaffolding molecules necessary for proper synaptogenesis. Interestingly, it was found that genetic deletion of NR3A accelerates glutamatergic synaptic transmission, as measured by AMPAR-mediated postsynaptic currents recorded in hippocampal CA1. Consistent, the deletion of NR3A accelerates the expression of the glutamate receptor subunits NR1, NR2A, and GluR1 sugesting that glutamatergic synapse maturation is critically dependent upon activation of NMDA-type glutamate receptors (Henson et al., 2012).
General role in biology: The development of neuronal networks can be distinguished into two phases: an early ‘establishment’ phase of neuronal connections, where activity-dependent and independent mechanisms could operate, and a later ‘maintenance’ phase, which appears to be controlled by neuronal activity (Yuste and Sur, 1999). These neuronal networks facilitate information flow that is necessary to produce complex behaviors, including learning and memory (Mayford et al., 2012).
How it is Measured or Detected
Methods that have been previously reviewed and approved by a recognized authority should be included in the Overview section above. All other methods, including those well established in the published literature, should be described here. Consider the following criteria when describing each method: 1. Is the assay fit for purpose? 2. Is the assay directly or indirectly (i.e. a surrogate) related to a key event relevant to the final adverse effect in question? 3. Is the assay repeatable? 4. Is the assay reproducible?
In vivo: The recording of brain activity by using electroencephalography (EEG), electrocorticography (ECoG) and local field potentials (LFP) assists towards the collection of signals generated by multiple neuronal cell networks. Advances in computer technology have allowed quantification of the EEG and expansion of quantitative EEG (qEEG) analysis providing a sensitive tool for time-course studies of different compounds acting on neuronal networks' function (Binienda et al., 2011). The number of excitatory or inhibitory synapses can be functionally studied at an electrophysiological level by examining the contribution of glutamatergic and GABAergic synaptic inputs. The number of them can be determined by variably clamping the membrane potential and recording excitatory and inhibitory postsynaptic currents (EPSCs or IPSCs) (Liu, 2004).
In vitro: Microelectrode array (MEA) recordings are also used to measure electrical activity in cultured neurons (Keefer et al., 2001, Gramowski et al., 2000; Gopal, 2003; Johnstone et al., 2010). MEAs can be applied in high throughput platforms to facilitate screening of numerous chemical compounds (McConnell et al., 2012). Using selective agonists and antagonists of different classes of receptors their response can be evaluated in a quantitative manner (Novellino et al., 2011; Hogberg et al., 2011).
Patch clamping technique can also be used to measure neuronal network activity.In some cases, if required, planar patch clamping technique can also be used to measure neuronal networks activity (e.g., Bosca et al., 2014).
References
Adams MM, Flagg RA, Gore AC., Perinatal changes in hypothalamic N-methyl-D-aspartate receptors and their relationship to gonadotropin-releasing hormone neurons. Endocrinology. 1999 May;140(5):2288-96.
Binienda ZK, Beaudoin MA, Thorn BT, Ali SF. (2011) Analysis of electrical brain waves in neurotoxicology: γ-hydroxybutyrate. Curr Neuropharmacol. 9: 236-239.
Bosca, A., M. Martina, and C. Py (2014) Planar patch clamp for neuronal networks--considerations and future perspectives. Methods Mol Biol, 2014. 1183: p. 93-113.
Erecinska M, Cherian S, Silver IA. (2004) Energy metabolism in mammalian brain during development. Prog Neurobiol. 73: 397-445.
Gambrill AC, Barria A. NMDA receptor subunit composition controls synaptogenesis and synapse stabilization. Proc Natl Acad Sci U S A. 2011:108(14):5855-60.
Gopal K. (2003) Neurotoxic effects of mercury on auditory cortex networks growing on microelectrode arrays: a preliminary analysis. Neurotoxicol Teratol. 25: 69-76.
Gramowski A, Schiffmann D, Gross GW. (2000) Quantification of acute neurotoxic effects of trimethyltin using neuronal networks cultures on microelectrode arrays. Neurotoxicology 21: 331-342.
Henson MA, Larsen RS, Lawson SN, Pérez-Otaño I, Nakanishi N, Lipton SA, Philpot BD. (2012) Genetic deletion of NR3A accelerates glutamatergic synapse maturation. PLoS One. 7(8).
Hogberg HT, Sobanski T, Novellino A, Whelan M, Weiss DG, Bal-Price AK. (2011) Application of micro-electrode arrays (MEAs) as an emerging technology for developmental neurotoxicity: evaluation of domoic acid-induced effects in primary cultures of rat cortical neurons. Neurotoxicology 32: 158-168.
Johnstone AFM, Gross GW, Weiss D, Schroeder O, Shafer TJ. (2010) Use of microelectrode arrays for neurotoxicity testing in the 21st century Neurotoxicology 31: 331-350.
Keefer E, Norton S, Boyle N, Talesa V, Gross G. (2001) Acute toxicity screening of novel AChE inhibitors using neuronal networks on microelectrode arrays. Neurotoxicology 22: 3-12.
Liu G. (2004) Local structural balance and functional interaction of excitatory and inhibitory synapses in hippocampal dendrites. Nat Neurosci. 7: 373-379.
Mayford M, Siegelbaum SA, Kandel ER. (2012) Synapses and memory storage. Cold Spring Harb Perspect Biol. 4. pii: a005751.
McConnell ER, McClain MA, Ross J, LeFew WR, Shafer TJ. (2012) Evaluation of multi-well microelectrode arrays for neurotoxicity screening using a chemical training set. Neurotoxicology 33: 1048-1057.
Novellino A, Scelfo B, Palosaari T, Price A, Sobanski T, Shafer TJ, Johnstone AF, Gross GW, Gramowski A, Schroeder O, Jügelt K, Chiappalone M, Benfenati F, Martinoia S, Tedesco MT, Defranchi E, D'Angelo P, Whelan M. (2011) Development of micro-electrode array based tests for neurotoxicity: assessment of interlaboratory reproducibility with neuroactive chemicals. Front Neuroeng. 4: 4.
Yuste R, Peinado A, Katz LC. (1992) Neuronal domains in developing neocortex. Science 257: 665-669.
Event: 2108: Altered, white brain matter
Short Name: Altered, white brain matter
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Tissue |
List of Adverse Outcomes in this AOP
Event: 341: Impairment, Learning and memory
Short Name: Impairment, Learning and memory
Key Event Component
| Process | Object | Action |
|---|---|---|
| learning | decreased | |
| memory | decreased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Individual |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| human | Homo sapiens | High | NCBI |
| rat | Rattus norvegicus | High | NCBI |
| fruit fly | Drosophila melanogaster | High | NCBI |
| zebrafish | Danio rerio | High | NCBI |
| gastropods | Physa heterostropha | High | NCBI |
| mouse | Mus musculus | High | NCBI |
| Life Stage | Evidence |
|---|---|
| During brain development | High |
| Adult, reproductively mature | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
Basic forms of learning behavior such as habituation have been found in many taxa from worms to humans (Alexander, 1990). More complex cognitive processes such as executive function likely reside only in higher mammalian species such as non-human primates and humans. Recently, larval zebrafish has also been suggested as a model for the study of learning and memory (Roberts et al., 2013).
Life stage applicability: This key event is applicable to various life stages such as during brain development and maturity (Hladik & Tapio, 2016).
Sex applicability: This key event is not sex specific (Cekanaviciute et al., 2018), although sex-dependent cognitive outcomes have been recently ; Parihar et al., 2020).
Evidence for perturbation by a prototypic stressor: Current literature provides ample evidence of impaired learning and memory being induced by ionizing radiation (Cekanaviciute et al., 2018; Hladik & Tapio, 2016).
Key Event Description
(Adapted from KE: 341 - in blue)
Learning can be defined as the process by which new information is acquired to establish knowledge by systematic study or by trial and error (Ono, 2009). Two types of learning are considered in neurobehavioral studies: a) associative learning and b) non- associative learning. Associative learning is based on making associations between different events. In associative learning, a subject learns the relationship among two different stimuli or between the stimulus and the subject’s behavior. On the other hand, non-associative learning can be defined as an alteration in the behavioral response that occurs over time in response to a single type of stimulus. Habituation and sensitization are some examples of non-associative learning.
The memory formation requires acquisition, retention and retrieval of information in the brain, which is characterized by the non- conscious recall of information (Ono, 2009). There are three main categories of memory, including sensory memory, short-term or working memory (up to a few hours) and long-term memory (up to several days or even much longer).
Learning and memory depend upon the coordinated action of different brain regions and neurotransmitter systems constituting functionally integrated neural networks (D’Hooge and DeDeyn, 2001). Among the many brain areas engaged in the acquisition of, or retrieval of, a learned event, the hippocampal-based memory systems have received the most study. For example, the hippocampus has been shown to be critical for spatial-temporal memory, visio-spatial memory, verbal and narrative memory, and episodic and autobiographical memory (Burgess et al., 2000; Vorhees and Williams, 2014). However, there is substantial evidence that fundamental learning and memory functions are not mediated by the hippocampus alone but require a network that includes, in addition to the hippocampus, anterior thalamic nuclei, mammillary bodies cortex, cerebellum and basal ganglia (Aggleton and Brown, 1999; Doya, 2000; Mitchell et al., 2002, Toscano and Guilarte, 2005; Gilbert et al., 2006, 2016). Thus, damage to variety of
brain structures can potentially lead to impairment of learning and memory. The main learning areas and pathways are similar in rodents and primates, including man (Eichenbaum, 2000; Stanton and Spear, 1990). While the prefrontal cortex and frontostriatal neural circuits have been identified as the primary sites of higher-order cognition in vertebrates, invertebrates utilize paired mushroom bodies, shown to contain ~300,000 neurons in honey bees (Menzel, 2012; Puig et al., 2014).
For the purposes of this KE (AO), impaired learning and memory is defined as an organism’s inability to establish new associative or non-associative relationships, or sensory, short-term or long-term memories which can be measured using different behavioral tests described below.
How it is Measured or Detected
In laboratory animals: in rodents, a variety of tests of learning and memory have been used to probe the integrity of hippocampal function. These include tests of spatial learning like the radial arm maze (RAM), the Barnes maze, Hebb-Williams maze, passive avoidance and Spontaneous alternation and most commonly, the Morris water maze (MWM). Test of novelty such as novel object recognition, and fear based context learning are also sensitive to hippocampal disruption. Finally, trace fear conditioning which incorporates a temporal component upon traditional amygdala-based fear learning engages the hippocampus. A brief description of these tasks follows.
RAM, Barnes, MWM, Hebb-Williams maze are examples of spatial tasks, animals are required to learn the location of a food reward (RAM); an escape hole to enter a preferred dark tunnel from a brightly lit open field area (Barnes maze), or a hidden platform submerged below the surface of the water in a large tank of water (MWM) (Vorhees and Williams, 2014). The Hebb- Williams maze measures an animal’s problem solving abilities by providing no spatial cues to find the target (Pritchett & Mulder, 2004).
Novel Object recognition. This is a simpler task that can be used to probe recognition memory. Two objects are presented to animal in an open field on trial 1, and these are explored. On trial 2, one object is replaced with a novel object and time spent interacting with the novel object is taken evidence of memory retention – I have seen one of these objects before, but not this one (Cohen and Stackman, 2015).
Contextual Fear conditioning is a hippocampal based learning task in which animals are placed in a novel environment and allowed to explore for several minutes before delivery of an aversive stimulus, typically a mild foot shock. Upon reintroduction to this same environment in the future (typically 24-48 hours after original training), animals will limit their exploration, the context of this chamber being associated with an aversive event. The degree of suppression of activity after training is taken as evidence of retention, i.e., memory (Curzon et al., 2009).
Trace fear conditioning. Standard fear conditioning paradigms require animals to make an association between a neutral conditioning stimulus (CS, a light or a tone) and an aversive stimulus (US, a footshock). The unconditioned response (CR) that is elicited upon delivery of the footshock US is freezing behavior. With repetition of CS/US delivery, the previously neutral stimulus comes to elicit the freezing response. This type of learning is dependent on the amygdala, a brain region associated with, but distinct from the hippocampus. Introducing a brief delay between presentation of the neutral CS and the aversive US, a trace period, requires the engagement of the amygdala and the hippocampus (Shors et al., 2001).
Operant Responding. Performance on operant responding reflects the cortex’ ability to organize processes (Rabin et al., 2002).
In humans: A variety of standardized learning and memory tests have been developed for human neuropsychological testing, including children (Rohlman et al., 2008). These include episodic autobiographical memory, perceptual motor tests, short and long term memory tests, working memory tasks, word pair recognition memory; object location recognition memory. Some have been incorporated in general tests of intelligence (IQ) such as the Wechsler Adult Intelligence Scale (WAIS) and the Wechsler.
Modifications have been made and norms developed for incorporating of tests of learning and memory in children. Examples of some of these tests include:
Rey Osterieth Complex Figure test (RCFT) which probes a variety of functions including as visuospatial abilities, memory, attention, planning, and working memory (Shin et al., 2006).
Children’s Auditory Verbal Learning Test (CAVLT) is a free recall of presented word lists that yields measures of Immediate Memory Span, Level of Learning, Immediate Recall, Delayed Recall, Recognition Accuracy, and Total Intrusions. (Lezak 1994; Talley, 1986).
Continuous Visual Memory Test (CVMT) measures visual learning and memory. It is a free recall of presented pictures/objects rather than words but that yields similar measures of Immediate Memory Span, Level of Learning, Immediate Recall, Delayed Recall, Recognition Accuracy, and Total Intrusions. (Lezak, 1984; 1994).
Story Recall from Wechsler Memory Scale (WMS) Logical Memory Test Battery, a standardized neurospychological test designed to measure memory functions (Lezak, 1994; Talley, 1986).
Autobiographical memory (AM) is the recollection of specific personal events in a multifaceted higher order cognitive process. It includes episodic memory- remembering of past events specific in time and place, in contrast to semantic autobiographical memory is the recollection of personal facts, traits, and general knowledge. Episodic AM is associated with greater activation of the hippocampus and a later and more gradual developmental trajectory. Absence of episodic memory in early life (infantile amnesia) is thought to reflect immature hippocampal function (Herold et al., 2015; Fivush, 2011).
Staged Autobiographical Memory Task. In this version of the AM test, children participate in a staged event involving a tour of the hospital, perform a series of tasks (counting footprints in the hall, identifying objects in wall display, buy lunch, watched a video). It is designed to contain unique event happenings, place, time, visual/sensory/perceptual details. Four to five months later, interviews are conducted using Children’s Autobiographical Interview and scored according to standardized scheme (Willoughby et al., 2014).
Attentional set-shifting (ATSET) task. Measures the ability to relearn cues over various schedules of reinforcement (Heisler et al., 2015).
In Honey Bees: For over 50 years an assay for evaluating olfactory conditioning of the proboscis extension reflex (PER) has been used as a reliable method for evaluating appetitive learning and memory in honey bees (Guirfa and Sandoz, 2012; LaLone et al., 2017). These experiments pair a conditioned stimulus (e.g., an odor) with an unconditioned stimulus (e.g., sucrose) provided immediately afterward, which elicits the proboscis extension (Menzel, 2012). After conditioning, the odor alone will lead to the conditioned PER. This methodology has aided in the elucidation of five types of olfactory memory phases in honey bee, which include early short-term memory, late short-term memory, mid-term memory, early long-term memory, and late long-term memory (Guirfa and Sandoz, 2012). These phases are dependent on the type of conditioned stimulus, the intensity of the unconditioned stimulus, the number of conditioning trials, and the time between trials. Where formation of short-term memory occurs minutes after conditioning and decays within minutes, memory consolidation or stabilization of a memory trace after initial acquisition leads to
mid-term memory, which lasts 1 d and is characterized by activity of the cAMP-dependent PKA (Guirfa and Sandoz, 2012). Multiple conditioning trials increase the duration of the memory after learning and coincide with increased Ca2+-calmodulin-dependent PKC activity (Guirfa and Sandoz, 2012). Early long-term memory, where a conditioned response can be evoked days to weeks after conditioning requires translation of existing mRNA, whereas late long-term memory requires de novo gene transcription and can last for weeks (Guirfa andSandoz, 2012)."
Regulatory Significance of the AO
A prime example of impairments in learning and memory as the adverse outcome for regulatory action is developmental lead exposure and IQ function in children (Bellinger, 2012). Most methods are well established in the published literature and many have been engaged to evaluate the effects of developmental thyroid disruption. The US EPA and OECD Developmental Neurotoxicity (DNT) Guidelines (OCSPP 870.6300 or OECD TG 426) as well as OECD TG 443 (OECD, 2018) both require testing of learning and memory (USEPA, 1998; OECD, 2007) advising to use the following tests passive avoidance, delayed-matching-to-position for the adult rat and for the infant rat, olfactory conditioning, Morris water maze, Biel or Cincinnati maze, radial arm maze, T-maze, and acquisition and retention of schedule-controlled behavior. These DNT Guidelines have been deemed valid to identify developmental neurotoxicity and adverse neurodevelopmental outcomes (Makris et al., 2009).
Also, in the frame of the OECD GD 43 (2008) on reproductive toxicity, learning and memory testing may have potential to be applied in the context of developmental neurotoxicity studies. However, many of the learning and memory tasks used in guideline studies may not readily detect subtle impairments in cognitive function associated with modest degrees of developmental thyroid disruption (Gilbert et al., 2012).
References
Aggleton JP, Brown MW. (1999) Episodic memory, amnesia, and the hippocampal-anterior thalamic axis. Behav Brain Sci. 22: 425- 489.
Alexander RD (1990) Epigenetic rules and Darwinian algorithms: The adaptive study of learning and development. Ethology and Sociobiology 11:241-303.
Bellinger DC (2012) A strategy for comparing the contributions of environmental chemicals and other risk factors to neurodevelopment of children. Environ Health Perspect 120:501-507.
Burgess N (2002) The hippocampus, space, and viewpoints in episodic memory. Q J Exp Psychol A 55:1057-1080. Cohen, SJ and Stackman, RW. (2015). Assessing rodent hippocampal involvement in the novel object recognition task. A review. Behav. Brain Res. 285: 105-1176.
Cekanaviciute, E., S. Rosi and S. Costes. (2018), "Central Nervous System Responses to Simulated Galactic Cosmic Rays", International Journal of Molecular Sciences, Vol. 19/11, Multidisciplinary Digital Publishing Institute (MDPI) AG, Basel, https://doi.org/10.3390/ijms19113669.
Cohen, SJ and Stackman, RW. (2015). Assessing rodent hippocampal involvement in the novel object recognition task. A review. Behav. Brain Res. 285: 105-1176.
Curzon P, Rustay NR, Browman KE. Cued and Contextual Fear Conditioning for Rodents. In: Buccafusco JJ, editor. Methods of Behavior Analysis in Neuroscience. 2nd edition. Boca Raton (FL): CRC Press/Taylor & Francis; 2009.
D'Hooge R, De Deyn PP (2001) Applications of the Morris water maze in the study of learning and memory. Brain Res Brain Res Rev 36:60-90.
Doya K. (2000) Complementary roles of basal ganglia and cerebellum in learning and motor control. Curr Opin Neurobiol. 10: 732- 739.
Eichenbaum H (2000) A cortical-hippocampal system for declarative memory. Nat Rev Neurosci 1:41-50. Fivush R. The development of autobiographical memory. Annu Rev Psychol. 2011;62:559-82.
Gilbert ME, Sanchez-Huerta K, Wood C (2016) Mild Thyroid Hormone Insufficiency During Development Compromises Activity- Dependent Neuroplasticity in the Hippocampus of Adult Male Rats. Endocrinology 157:774-787.
Gilbert ME, Rovet J, Chen Z, Koibuchi N. (2012) Developmental thyroid hormone disruption: prevalence, environmental contaminants and neurodevelopmental consequences. Neurotoxicology 33: 842-52.
Gilbert ME, Sui L (2006) Dose-dependent reductions in spatial learning and synaptic function in the dentate gyrus of adult rats following developmental thyroid hormone insufficiency. Brain Res 1069:10-22.
Guirfa, M., Sandoz, J.C., 2012. Invertebrate learning and memory: fifty years of olfactory conditioning of the proboscis extension response in honeybees. Learn. Mem. 19 (2),
54–66.
Herold, C, Lässer, MM, Schmid, LA, Seidl, U, Kong, L, Fellhauer, I, Thomann,PA, Essig, M and Schröder, J. (2015). Neuropsychology, Autobiographical Memory, and Hippocampal Volume in “Younger” and “Older” Patients with Chronic Schizophrenia. Front. Psychiatry, 6: 53.
Hladik, D. and S. Tapio. (2016), "Effects of ionizing radiation on the mammalian brain", Mutation Research/Reviews in Mutation Research, Vol. 770, Elsevier B. b., Amsterdam, https://doi.org/10.1016/j.mrrev.2016.08.003.
Heisler, J. M. et al. (2015), "The Attentional Set Shifting Task: A Measure of Cognitive Flexibility in Mice", Journal of Visualized Experiments, 96, JoVe, Cambridge, https://doi.org/10.3791/51944.
LaLone, C.A., Villeneuve, D.L., Wu-Smart, J., Milsk, R.Y., Sappington, K., Garber, K.V., Housenger, J. and Ankley, G.T., 2017. Weight of evidence evaluation of a network of adverse outcome pathways linking activation of the nicotinic acetylcholine receptor in honey bees to colony death. STOTEN. 584-585, 751-775.
Lezak MD (1984) Neuropsychological assessment in behavioral toxicology--developing techniques and interpretative issues. Scand J Work Environ Health 10 Suppl 1:25-29.
Lezak MD (1994) Domains of behavior from a neuropsychological perspective: the whole story. Nebr Symp Motiv 41:23-55.
Makris SL, Raffaele K, Allen S, Bowers WJ, Hass U, Alleva E, Calamandrei G, Sheets L, Amcoff P, Delrue N, Crofton KM. (2009) A retrospective performance assessment of the developmental neurotoxicity study in support of OECD test guideline 426. Environ Health Perspect. Jan;117(1):17-25.
Menzel, R., 2012. The honeybee as a model for understanding the basis of cognition. Nat. Rev. Neurosci. 13 (11), 758–768.
Mitchell AS, Dalrymple-Alford JC, Christie MA. (2002) Spatial working memory and the brainstem cholinergic innervation to the anterior thalamus. J Neurosci. 22: 1922-1928.
OECD. 2007. OECD guidelines for the testing of chemicals/ section 4: Health effects. Test no. 426: Developmental neurotoxicity study. www.Oecd.Org/dataoecd/20/52/37622194.Pdf [accessed may 21, 2012].
OECD (2008) Nr 43 GUIDANCE DOCUMENT ON MAMMALIAN REPRODUCTIVE TOXICITY TESTING AND ASSESSMENT. ENV/JM/MONO(2008)16
Ono T. (2009) Learning and Memory. Encyclopedia of neuroscience. M D. Binder, N. Hirokawa and U. Windhorst (Eds). Springer- Verlag GmbH Berlin Heidelberg. pp 2129-2137.
Parihar, V. K. et al. (2020), "Sex-Specific Cognitive Deficits Following Space Radiation Exposure", Frontiers in Behavioral Neuroscience, Vol. 14, https://doi.org/10.3389/fnbeh.2020.535885.
Pritchett, K. and G. Mulder. (2004), "Hebb-Williams mazes.", Contemporary topics in laboratory animal science, Vol. 43/5, http://www.ncbi.nlm.nih.gov/pubmed/15461441.
Puig, M.V., Antzoulatos, E.G., Miller, E.K., 2014. Prefrontal dopamine in associative learning and memory. Neuroscience 282, 217– 229.
Rabin, B. M. et al. (2002), "Effects of Exposure to 56Fe Particles or Protons on Fixed-ratio Operant Responding in Rats", Journal of Radiation Research, Vol. 43/S, https://doi.org/10.1269/jrr.43.S225.
Roberts AC, Bill BR, Glanzman DL. (2013) Learning and memory in zebrafish larvae. Front Neural Circuits 7: 126.
Rohlman DS, Lucchini R, Anger WK, Bellinger DC, van Thriel C. (2008) Neurobehavioral testing in human risk assessment. Neurotoxicology. 29: 556-567.
Shin, MS, Park, SY, Park, SR, Oeol, SH and Kwon, JS. (2006). Clinical and empirical applications of the Rey-Osterieth complex figure test. Nature Protocols, 1: 892-899.
Shors TJ, Miesegaes G, Beylin A, Zhao M, Rydel T, Gould E (2001) Neurogenesis in the adult is involved in the formation of trace memories. Nature 410:372-376.
Stanton ME, Spear LP (1990) Workshop on the qualitative and quantitative comparability of human and animal developmental neurotoxicity, Work Group I report: comparability of measures of developmental neurotoxicity in humans and laboratory animals. Neurotoxicol Teratol 12:261-267.
Talley, JL. (1986). Memory in learning disabled children: Digit span and eh Rey Auditory verbal learning test. Archives of Clinical Neuropsychology, Elseiver.
Toscano CD, Guilarte TR. (2005) Lead neurotoxicity: From exposure to molecular effects. Brain Res Rev. 49: 529-554.
U.S.EPA. 1998. Health effects guidelines OPPTS 870.6300 developmental neurotoxicity study. EPA Document 712-C-98-239.Office of Prevention Pesticides and Toxic Substances.
Vorhees CV, Williams MT (2014) Assessing spatial learning and memory in rodents. ILAR J 55:310-332.
Willoughby KA, McAndrews MP, Rovet JF. Accuracy of episodic autobiographical memory in children with early thyroid hormone deficiency using a staged event. Dev Cogn Neurosci. 2014 Jul;9:1-11.
Event: 2231: Locomotor function. Impaired
Short Name: Impaired locomotor function
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Population |
Appendix 2
List of Key Event Relationships in the AOP
List of Adjacent Key Event Relationships
Relationship: 3651: Inhibition, monocarboxylate transporter 8 (MCT8) leads to Decreased brain T4/T3
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3652: Inhibition, OATP1C1 leads to Decreased brain T4/T3
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3653: Inhibition, Deiodinase 2 leads to Decreased brain T4/T3
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3654: Decreased brain T4/T3 leads to TR Antagnoism
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3655: TR Antagnoism leads to BDNF, Reduced
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 448: BDNF, Reduced leads to Synaptogenesis, Decreased
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Inhibition of Na+/I- symporter (NIS) leads to learning and memory impairment | non-adjacent | Moderate | Low |
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Evidence Supporting Applicability of this Relationship
| Life Stage | Evidence |
|---|---|
| During brain development | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
Empirical evidence comes from work with laboratory rodent-derived cells and brain slices, and rodent in vivo studies.
Key Event Relationship Description
Disruption of BDNF signaling (and other factors, such as NGF or Reelin, etc.) during brain development was shown to interfere with synaptogenesis in the hippocampus (Sanchez-Martin et al., 2013; Neal et al., 2010; Stansfiled et al., 2012). In the adult brain, BDNF is involved in synaptic plasticity (Lu et al., 2013; Leal et al., 2014), which is a fundamental process linked with learning and memory. Synaptic dysfunction is a key pathophysiological hallmark in neurodegenerative disorders, including Alzheimer's disease, and synaptic repair therapies based on the use of trophic factors, such as BDNF, are currently under consideration (Lu et al., 2013).
BDNF is released by the BDNF-producing neurons of the CNS and binds to Trk-B of the PV-interneurons, an interaction necessary for the subsequent developmental effects of this neurotrophin (Polleux et al., 2002; Jin et al., 2003; Rico et al., 2002; Aguado et al., 2003). BDNF promotes the morphological and neurochemical maturation of hippocampal and neocortical interneurons and promotes GABAergic synaptogenesis (Danglot et al., 2006; Hu and Russek, 2008).
BDNF plays an important role in axonal and dendritic differentiation during embryonic stages of neuronal development, as well as in the formation and maturation of dendritic spines during postnatal development (Chapleau et al., 2009). Recent studies have also implicated vesicular trafficking of BDNF via secretory vesicles, and both secretory and endosomal trafficking of vesicles containing synaptic proteins, such as neurotransmitter and neurotrophin receptors, in the regulation of axonal and dendritic differentiation, and in dendritic spine morphogenesis. Abnormalities in dendritic and synaptic structure are consistently observed in human neurodevelopmental disorders associated with mental retardation, as well as in mouse models of these disorders (Chapleau et al., 2009).
Evidence Supporting this KER
Biological PlausibilityBDNF, in addition to its pro-survival effects, has powerful synaptic effects, promoting synaptic transmission, synaptic plasticity and synaptogenesis (Lu et al., 2013; Sanchez-Martin et al., 2013; Neal et al., 2010; Stansfiled et al., 2012; Danglot et al., 2006; Hu and Russek, 2008). NMDAR activity has been linked to the signaling of the trans-synaptic neurotorophin BDNF (Neal et al., 2010).
Use of selective agonist or antagonist of BDNF receptor TrkB demonstrates the contribution of BDNF in synaptogenesis in adult-generated neurons in the rat dentate gyrus (Ambrogini et al., 2013). In this regard, exogenous application of BDNF significantly increased the number of functional synapses in culture (Vicario-Abejon et al., 1998; Marty et al., 2000), while blocking of BDNF with antibodies greatly reduced the formation of inhibitory synapses (Seil and Drake-Baumann, 2000). Similar results were described also in an in vivo study on mutant mice characterized by deletion of the trkB gene in cerebellar precursors (obtained by Wnt1-driven Cre--mediated recombination). TrkB mutant mice showed reduced amounts of GABAergic markers and develop reduced numbers of GABAergic boutons and synaptic specializations, whilst granule and Purkinje cell dendrites appeared normal and the former presented typical numbers of excitatory synapses. This study demonstrated that TrkB is essential to the development of GABAergic neurons and the regulation of synapse formation (Rico et al., 2002). BDNF is also a potent regulator of spontaneous neuronal activity in GABAergic neurons and interneurons, as shown in in embryonic (E18) hippocampal slices (Aguado et al., 2003), and plays a critical role in controlling the emergence, complexity and networking properties of spontaneous networks.
TH deficiency during the foetal and/or the neonatal period, apart from reducing synaptogenesis, can produce several other deleterious effects for neural growth and development (e.g., such as reduced synaptic connectivity, delayed myelination, disturbed neuronal migration, deranged axonal projections, and alterations in neurotransmitters' levels), possibly through decreased BDNF levels (Koromilas et al., 2010; Shafiee et al., 2016).
Empirical EvidenceSeveral studies (in vitro, ex vivo, and in vivo) have shown correlations between downregulation of BDNF signaling (e.g., in trasgenic animals, or upon treatment with K252a (a BDNF receptor inhibitor) or with an antibody anti-BDNF) and synaptogenesis (and synapses) decrease:
- Westerholz et al., 2013 In recent in vitro studies with rat T3-deficient cultures of cortical GABAergic PV+ interneurons, which are subject to BDNF regulation, it was shown that the number of synaptic boutons (i.e., presynaptic terminals containing the presynaptic marker synaptophysin) was reduced, an effect that was abolished after exogenous BDNF application. Additionally, inhibition of BDNF TrkB receptors by K252a in cultures containing T3 resulted also in decreased number of synaptic boutons, as in the T3-deprived cultures. These results indicate that BDNF signaling promotes the formation of synaptic boutons and that this function is mediated by THs (T3 and T4). Additionally, T3-related increase of spontaneous network activity was remarkably reduced after addition of K252a, and also upon inhibition of mTOR pathway (with rapamycin), a pathway known to control synaptogenesis (Buckmaster et al., 2009).
- Sato et al., 2007 This study on rat cultured hippocampal slices showed that beta-estradiol (E2) induced synaptogenesis between mossy fibers (one of the major inputs to cerebellum) and hippocampal CA3 neurons by enhancing BDNF release from dentate gyrus (DG) granule cells, by increasing the expression of PSD95, a postsynaptic marker. E2 effects on in hippocampal slice cultures and subregional neuron cultures were completely inhibited by blocking the BDNF receptor (TrkB) with K252a (200 nM) or by using a function-blocking antibody to BDNF (10 μg/ml), which inhibited the expression of PSD95 induced by E2. Both K252a and the antibody anti-BDNF elicited ~ 60-70% decrease of spine density and ~ 55% decrease of presynaptic sites in dentate gyrus granule cells (measured as number of puncta/neuron).
- Schjetnan and Escobar, 2012 In this study, intrahippocampal microinfusion of BDNF (3 µg/3 µl; 0.2 µl/min,) in adult rats modified the ability of the hippocampal mossy fiber pathway to present long-term potentiation (LTP, i.e., a persistent strengthening of synapses based on recent patterns of activity) by high frequency stimulation (HFS). This indicates that BDNF initiates the metaplastic mechanisms that modify the ability of the mossy fiber pathway to present LTP induced by subsequent HFS. On the contrary, microinfusion of K252a (administered in combination with BDNF: 3 μg of BDNF/3 μl of K252a 20 μM; 0.2 μl/min) blocked the functional and morphological effects produced by BDNF (shown by densitometric analysis on synaptic reorganization: ~ 30% reduction of the relative area of the dorsal hippocampus in the contralateral side of HFS, and ~ 70% reduction in the ipsilateral side of HFS, compared to BDNF administered alone), supporting the role of BDNF in the regulation of synaptic plasticity.
- Schildt et al., 2013 Using field potential recordings in CA3 of adult heterozygous BDNF knockout (BDNF+/-) mice, an impairment of NMDAR-independent mossy fiber (MF)-LTP (~ 50% decrease) was observed. Additionally, inhibition of TrkB/BDNF with K252a (slices preincubated for 3 hr with 100 nM), or with the selective BDNF scavenger TrkB-Fc (slices preincubated for 3 hr with 5 μg/ml), both inhibited MF-LTP to the same extent as observed in BDNF+/- mice (K252a: ~ 60% decrease vs control slices; TrkB-Fc: ~ 50% decrease vs control slices).
- Cortés et al., 2012 Adult male Sprague-Dawley rats were treated with 6-propyl-2-thiouracil (PTU, a TPO inhibitor) (0.05% in drinking water) for 20 days to induce hypothyroidism. PTU-treated rats showed decrease serum fT4 (~ 70% decrease vs control) and tT3 (~ 45% decrease vs control) levels, and increased TSH levels (~ 9.5-fold increase over control). The hippocampus of hypothyroid adult rats displayed increased apoptosis levels in neurons and astrocyte and reactive gliosis compared with controls. The glutamatergic synapses from the stratum radiatum of CA3 from hypothyroid rats, contained lower postsynaptic density (PSD) than control rats (~ 25% lower PSD than control). This observation was in agreement with a reduced content of NMDAR subunits (NR1 and NR2A/B subunits, both subunits: ~ 25% decrease vs control) at the PSD in hypothyroid animals. Additionally, the hippocampal amount of BDNF mRNA (assessed by in situ hybridization) was higher (~ 4.8-fold increase over control) of hypothyroid rats, while the content of TrkB protein (BDNF receptor) was reduced (~ 30% decrease vs control) at the PSD of the CA3 region of hypothyroid rats, compared with controls. Even though BDNF levels were increased, the decrease of BDNF receptor (TrkB) compromises the signalling pathway under BDNF control.
- Koibuchi et al., 2001 Here newborn mice were rendered hypothyroid by administering MMI (TPO inhibitor) and perchlorate (NIS inhibitor) in drinking water to their mothers. Neurotrophin-3 (NT-3) and BDNF gene expression was depressed in the perinatal hypothyroid cerebellum. Furthermore, the expression of retinoid-receptor-related orphan nuclear hormone receptor-alpha (ROR-alpha), an orphan nuclear receptor that plays critical roles in Purkinje cell development, was also decreased. Morphologically, disappearance of the external granule cell layer was retarded and arborization of Purkinje cell dendrite was decreased in hypothyroid rats. Dendritic arborization is used as readout for synapse formation, as post-synaptic side (synaptogenesis) is mainly located on dendrites.
- Aguado et al., 2003 BDNF overexpression in transgenic embryos raised the spontaneous activity of E18 hippocampal neurons, as shown by increased number of synapses (63% more synapses in the hippocampus of BDNF transgenic embryos than in controls), and increased spontaneous neuronal activity (2.3 times more active neurons than wild type embryos, and 36.3% greater rates of activation). Moreover, BDNF transgenic embryos had higher number of GABAergic interneuron synapses, as shown by higher GAD67 mRNA (by 3-fold) and K(+)/Cl(-) KCC2 mRNA expression (by 4.3-fold) (responsible for the conversion of GABA responses from depolarizing to inhibitory), without altering the expression of GABA and glutamate ionotropic receptors. These data indicate that BDNF controls both GABAergic pre- and postsynaptic sites.
KEs proceeding the AO (decreased cognition), such as "Reduced BDNF Release" and "Decreased synaptogenesis" are also common to the AOP 13, entitled "Chronic binding of antagonist to N-methyl-D-aspartate receptors (NMDARs) during brain development induces impairment of learning and memory abilities" (https://aopwiki.org/aops/13). In this AOP 13, data on lead (Pb) exposure, as a reference chemical, are reported. These studies do not refer to TH disruption; however, they provide empirical support for this KER (Reduced release of BDNF leads to decreased synaptogenesis).
Synaptic structural plasticity was shown to be modified by Pb treatment during early (pre-weaning) or late (post-weaning) brain development in rats exposed to 2 mM Pb in drinking water for 3 weeks (Xiao et al., 2014). An iron chelator (clioquinol) can rescue the Pb-induced impairment of synaptic plasticity in hippocampus (Chen et al., 2007), showing that Pb can affect synaptogenesis and synaptic plasticity. Primary hippocampal neurons obtained from ED18 rat pups and treated with Pb (1, 2 microM) for 5 days exhibited pre-synaptic deficits due to disruption of NMDAR-dependent BDNF signaling (Neal et al., 2010; Stansfield et al., 2012). A decrease in bdnf expression was observed in mouse embryonic stem cells differentiated into neurons, if they were exposed to Pb 0.1 microM throughout the whole differentiation process (Sanchez-Martin et al., 2013). Similar alterations in gene expression patterns of neural markers (synapsin 1), neurotrophins (bdnf), transcription factors and glutamate-related genes were found in mice, when their mothers were exposed to 0-3 ppm of Pb in drinking water from 8 weeks prior to mating, through gestation and until postnatal day 10 (Sanchez-Martin et al., 2013).
Uncertainties and InconsistenciesAlterations of BDNF signaling is probably not the only mechanism leading to impaired synaptogenesis and synaptic plasticity. Indeed NMDAR activity can also modulate nitric oxide (NO) signaling. Exogenous NO addition during Pb exposure results in complete recovery of whole-cell synaptophysin levels and partial recovery of synaptophysin and synaptobrevin in synapses in Pb-exposed neurons (Neal et al., 2012). In addition, in Wistar rats, the anti-oxidant and radical scavenger quercetin was able to relieve the impairment of synaptic plasticity induced by chronic Pb exposure (from parturition through adulthood (PND 60); 0.2% Pb in drinking water of mothers and post-weaning pups) (Hu et al., 2008), suggesting that oxidative stress can also interfere with synapse formation.
Additionally, while PTU (a TPO inhibitor) has been shown to decrease brain BDNF levels and expression in offspring born from PTU-treated rat dams (Shafiee et al. 2016; Chakraborty et al., 2012; Gilbert et al. 2016), in the study from Cortés and colleagues (Cortés et al., 2012), treatment of adult male Sprague-Dawley rats with PTU induced an increase in the amount of BDNF mRNA in the hippocampus, while the content of TrkB protein, the BDNF receptor, resulted reduced at the PSD of the CA3 region compared with controls. Treated rats presented also thinner PSD than control rats, and a reduced content of NMDAr subunits (NR1 and NR2A/B subunits) at the PSD. These indicate differential effects elicited by PTU (i.e., TPO inhibition) on BDNF expression/regulation comparing the adult vs foetal brain. However, even though BDNF levels were increased, the decrease of BDNF receptor (TrkB) compromises the signalling pathway under BDNF control.
Results variability from study to study is due to different experimental study designs, accounting for differences in brain development stages (PND vs adult), times of exposures to chemicals, and regional brain differences.
References
Aguado F, Carmona MA, Pozas E, Aguiló A, Martínez-Guijarro FJ, Alcantara S, Borrell V, Yuste R, Ibañez CF, SorianoE. (2003). BDNF regulates spontaneous correlated activity at early developmental stages by increasing synaptogenesis and expression of the K+/Cl–co-transporter KCC2. Development 130:1267-1280.
Ambrogini P, Lattanzi D, Ciuffoli S, Betti M, Fanelli M, Cuppini R. (2013). Physical exercise and environment exploration affect synaptogenesis in adult-generated neurons in the rat dentate gyrus: possible role of BDNF. Brain Res 1534: 1-12.
Buckmaster PS, Ingram EA, Wen X. (2009). Inhibition of the mammalian target of rapamycin signaling pathway suppresses dentate granule cell axon sprouting in a rodent model of temporal lobe epilepsy. J Neurosci. Jun 24; 29(25):8259-69.
Chakraborty G, Magagna-Poveda A, Parratt C, Umans JG, MacLusky NJ, Scharfman HE. (2012). Reduced hippocampal brain-derived neurotrophic factor (BDNF) in neonatal rats after prenatal exposure to propylthiouracil (PTU). Endocrinology 153:1311–1316.
Chapleau CA, Larimore JL, Theibert A, Pozzo-Miller L. (2009). Modulation of dendritic spine development and plasticity by BDNF and vesicular trafficking: fundamental roles in neurodevelopmental disorders associated with mental retardation and autism. J Neurodev Disord;1:185–196.
Chen WH, Wang M, Yu SS, Su L, Zhu DM, She JQ, et al. (2007). Clioquinol and vitamin B12 (cobalamin) synergistically rescue the lead-induced impairments of synaptic plasticity in hippocampal dentate gyrus area of the anesthetized rats in vivo. Neuroscience 147(3): 853-864.
Cortés C, Eugenin E, Aliaga E, Carreño LJ, Bueno SM, Gonzalez PA, Gayol S, Naranjo D, Noches V, Marassi MP, Rosenthal D, Jadue C, Ibarra P, Keitel C, Wohllk N, Court F, Kalergis AM, Riedel CA. (2012). Hypothyroidism in the adult rat causes incremental changes in brain-derived neurotrophic factor, neuronal and astrocyte apoptosis, gliosis, and deterioration of postsynaptic density. Thyroid. Sep;22(9):951-63.
Danglot L, Triller A, Marty S. (2006). The development of hippocampal interneurons in rodents. Hippocampus. 16:1032-1060.
Gilbert ME, Sanchez-Huerta K, Wood C. (2016). Mild Thyroid Hormone Insufficiency During Development Compromises Activity-Dependent Neuroplasticity in the Hippocampus of Adult Male Rats. Endocrinology, Feb;157(2):774-87
Hu Y, Russek SJ. (2008). BDNF and the diseased nervous system: a delicate balance between adaptive and pathological processes of gene regulation. J Neurochem. 105:1-17.
Hu P, Wang M, Chen WH, Liu J, Chen L, Yin ST, et al. (2008). Quercetin relieves chronic lead exposure-induced impairment of synaptic plasticity in rat dentate gyrus in vivo. Naunyn Schmiedebergs Arch Pharmacol. Jul;378(1):43-51.
Jin X, Hu H, Mathers PH, Agmon A. (2003). Brain-derived neurotrophic factor mediates activity-dependent dendritic growth in nonpyramidal neocortical interneurons in developing organotypic cultures. J Neurosci 23:5662–5673.
Koibuchi N, Yamaoka S, Chin WW. (2001). Effect of altered thyroid status on neurotrophin gene expression during postnatal development of the mouse cerebellum. Thyroid 11:205–210.
Koromilas C, Liapi C, Schulpis KH, Kalafatakis K, Zarros A, Tsakiris S. Structural and functional alterations in the hippocampus due to hypothyroidism. Metab Brain Dis. 2010 Sep;25(3):339-54.
Leal G, Comprido D, Duarte CB. (2014). BDNF-induced local protein synthesis and synaptic plasticity. Neuropharmacology 76 Pt C: 639-656.
Lu B, Nagappan G, Guan X, Nathan PJ, Wren P. (2013). BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat Rev Neurosci 14(6): 401-416.
Marty S, Wehrle R, Sotelo C. (2000). Neuronal activity and brain-derived neurotrophic factor regulate the density of inhibitory synapses in organotypic slice cultures of postnatal hippocampus. J Neurosci 20: 8087–8095.
Neal AP, Stansfield KH, Worley PF, Thompson RE, Guilarte TR. (2010). Lead exposure during synaptogenesis alters vesicular proteins and impairs vesicular release: potential role of NMDA receptor-dependent BDNF signaling. Toxicol Sci 116(1): 249-263.
Neal AP, Stansfield KH, Guilarte TR. (2012). Enhanced nitric oxide production during lead (Pb(2)(+)) exposure recovers protein expression but not presynaptic localization of synaptic proteins in developing hippocampal neurons. Brain Res 1439: 88-95.
Polleux F, Whitford KL, Dijkhuizen PA, Vitalis T, Ghosh A. (2002). Control of cortical interneuron migration by neurotrophins and PI3-kinase signaling. Development 129:3147–60.
Rico B, Xu B, Reichardt LF. (2002). TrkB receptor signaling is required for establishment of GABAergic synapses in the cerebellum. Nat Neurosci 5:225–233.
Sanchez-Martin FJ, Fan Y, Lindquist DM, Xia Y, Puga A. (2013). Lead induces similar gene expression changes in brains of gestationally exposed adult mice and in neurons differentiated from mouse embryonic stem cells. PLoS One 8(11): e80558.
Sato K, Akaishi T, Matsuki N, Ohno Y, Nakazawa K. (2007). beta-Estradiol induces synaptogenesis in the hippocampus by enhancing brain-derived neurotrophic factor release from dentate gyrus granule cells. Brain Res. May 30;1150:108-20.
Schildt S, Endres T, Lessmann V, Edelmann E. (2013). Acute and chronic interference with BDNF/TrkB-signaling impair LTP selectively at mossy fiber synapses in the CA3 region of mouse hippocampus. Neuropharmacology. Aug;71:247-54.
Schjetnan AG, Escobar ML. (2012). In vivo BDNF modulation of hippocampal mossy fiber plasticity induced by high frequency stimulation. Hippocampus. Jan;22(1):1-8.
Seil FJ, Drake-Baumann R. (2000). TrkB receptor ligands promote activity-dependent inhibitory synaptogenesis. J Neurosci 20: 5367–73.
Shafiee SM, Vafaei AA, Rashidy-Pour A. (2016). Effects of maternal hypothyroidism during pregnancy on learning, memory and hippocampal BDNF in rat pups: Beneficial effects of exercise. Neuroscience. Aug 4;329:151-61.
Stansfield KH, Pilsner JR, Lu Q, Wright RO, Guilarte TR. (2012). Dysregulation of BDNF-TrkB signaling in developing hippocampal neurons by Pb(2+): implications for an environmental basis of neurodevelopmental disorders. Toxicol Sci 127(1): 277-295.
Vicario-Abejon C, Collin C, McKay RD, Segal M. (1998). Neurotrophins induce formation of functional excitatory and inhibitory synapses between cultured hippocampal neurons. J Neurosci 18:7256–71
Westerholz S, de Lima AD, Voigt T. (2013). Thyroid hormone-dependent development of early cortical networks: temporal specificity and the contribution of trkB and mTOR pathways. Front Cell Neurosci 7:121.
Xiao Y, Fu H, Han X, Hu X, Gu H, Chen Y, et al. (2014). Role of synaptic structural plasticity in impairments of spatial learning and memory induced by developmental lead exposure in Wistar rats. PLoS One 9(12): e115556.
Relationship: 3656: Synaptogenesis, Decreased leads to neural network formation
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3657: neural network formation leads to Impaired locomotor function
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3659: neural network formation leads to Impairment, Learning and memory
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3660: TR Antagnoism leads to Cell cycle, disrupted
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3661: TR Antagnoism leads to Decreased, mbp expression in oligodendrocytes
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3662: Cell cycle, disrupted leads to Demyelination, increased
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3663: Decreased, mbp expression in oligodendrocytes leads to Demyelination, increased
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3664: Demyelination, increased leads to Impaired axonal insulation
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3665: Impaired axonal insulation leads to Neuronal network function, Decreased
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3666: Neuronal network function, Decreased leads to Altered, white brain matter
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3192: Altered, white brain matter leads to Impairment, Learning and memory
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Reduced oligodendrocyte differentiation during neurodevelopment leading to impaired learning and memory | adjacent | High | Not Specified |
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
Relationship: 3667: Altered, white brain matter leads to Impaired locomotor function
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | adjacent | Not Specified | Not Specified |
List of Non Adjacent Key Event Relationships
Relationship: 3658: Decreased brain T4/T3 leads to BDNF, Reduced
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Decreased thyroid hormone levels in the brain regulated via transport, metabolism and TR activation leading to decreased cognition and motor function | non-adjacent | Not Specified | Not Specified |